
2019年 第5卷 第1期
《工程(英文)》 >> 2019年 第5卷 第1期 doi: 10.1016/j.eng.2018.11.013
肠道微生态——探索中药临床疗效的新思路
aState Key Laboratory for Diagnosis and Treatment of Infectious Diseases, The First Affiliated Hospital, School of Medicine, Zhejiang University, Hangzhou 310003, China
bCollaborative Innovation Center for Diagnosis and Treatment of Infectious Diseases, Hangzhou 310003, China
下一篇 上一篇
摘要
自古以来,中药在疾病预防、症状缓解和健康改善等方面发挥了重要作用。然而,复杂的成分和尚未明确的机制阻碍了中药的广泛推广和应用。越来越多的研究表明,和人体健康息息相关的肠道微生态与中药的疗效有关,因此从肠道微生态的角度去探索中药可能是打开中药奥秘的金钥匙。肠道菌群主要通过以下4个生理途径中发挥作用:参与宿主代谢、调节系统免疫、维持胃肠道稳态以及影响脑功能和宿主行为。本文回顾了中药与慢性肝病、溃疡性结肠炎、肥胖和2型糖尿病等疾病之间的联系,从肠道微生态的角度阐明其潜在的机制。未来,我们需要进一步的研究和更完善的实验设计,以揭示中药与肠道菌群之间相互作用的具体机制,并为中药的创新研究提供新的思路。
图片

图1

图2
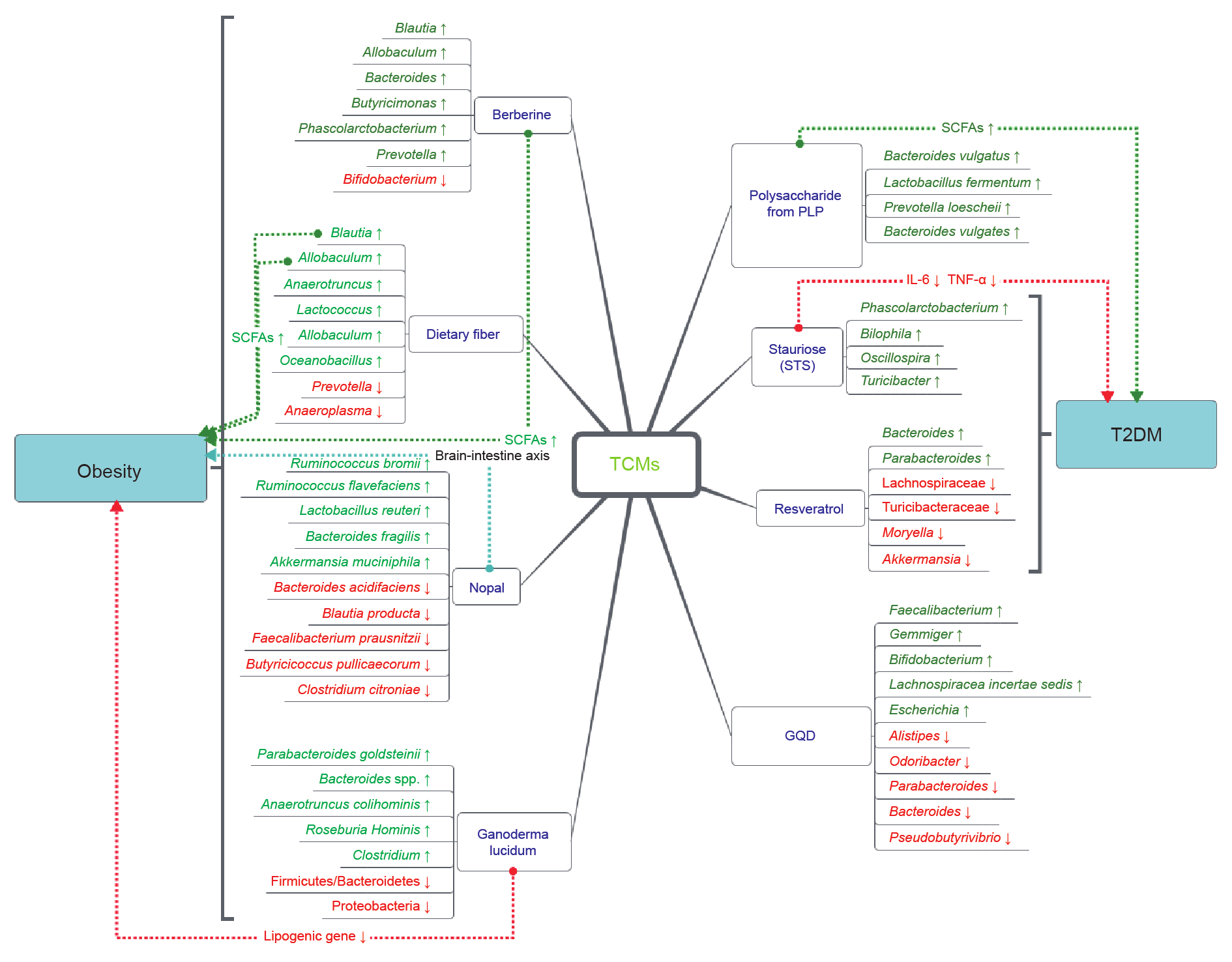
图3
参考文献
[ 1 ] Xu J, Chen HB, Li SL. Understanding the molecular mechanisms of the interplay between herbal medicines and gut microbiota. Med Res Rev 2017;37(5):1140–85. 链接1
[ 2 ] Wishart DS. Emerging applications of metabolomics in drug discovery and precision medicine. Nat Rev Drug Discov 2016;15(7):473–84. 链接1
[ 3 ] Harvey AL, Edrada-Ebel R, Quinn RJ. The re-emergence of natural products for drug discovery in the genomics era. Nat Rev Drug Discov 2015;14(2):111–29. 链接1
[ 4 ] Clemente JC, Ursell LK, Parfrey LW, Knight R. The impact of the gut microbiota on human health: an integrative view. Cell 2012;148(6):1258–70. 链接1
[ 5 ] Leung C, Rivera L, Furness JB, Angus PW. The role of the gut microbiota in NAFLD. Nat Rev Gastroenterol Hepatol 2016;13(7):412–25. 链接1
[ 6 ] Manichanh C, Borruel N, Casellas F, Guarner F. The gut microbiota in IBD. Nat Rev Gastroenterol Hepatol 2012;9(10):599–608. 链接1
[ 7 ] Cox AJ, West NP, Cripps AW. Obesity, inflammation, and the gut microbiota. Lancet Diabetes Endocrinol 2015;3(3):207–15. 链接1
[ 8 ] Knip M, Siljander H. The role of the intestinal microbiota in type 1 diabetes mellitus. Nat Rev Endocrinol 2016;12(3):154–67. 链接1
[ 9 ] Qin J, Li Y, Cai Z, Li S, Zhu J, Zhang F, et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012;490(7418):55–60. 链接1
[10] Collins SM. A role for the gut microbiota in IBS. Nat Rev Gastroenterol Hepatol 2014;11(8):497–505. 链接1
[11] Zitvogel L, Galluzzi L, Viaud S, Vétizou M, Daillère R, Merad M, et al. Cancer and the gut microbiota: an unexpected link. Sci Transl Med 2015;7 (271):271ps1. 链接1
[12] Li H, Zhou M, Zhao A, Jia W. Traditional Chinese medicine: balancing the gut ecosystem. Phytother Res 2009;23(9):1332–5. 链接1
[13] Chen F, Wen Q, Jiang J, Li HL, Tan YF, Li YH, et al. Could the gut microbiota reconcile the oral bioavailability conundrum of traditional herbs? J Ethnopharmacol 2016;179:253–64. 链接1
[14] O’Hara AM, Shanahan F. The gut flora as a forgotten organ. EMBO Rep 2006;7 (7):688–93. 链接1
[15] Bäckhed F, Ley RE, Sonnenburg JL, Peterson DA, Gordon JI. Host-bacterial mutualism in the human intestine. Science 2005;307(5717):1915–20. 链接1
[16] Nicholson JK, Holmes E, Kinross J, Burcelin R, Gibson G, Jia W, et al. Host-gut microbiota metabolic interactions. Science 2012;336(6086):1262–7. 链接1
[17] Hooper LV, Wong MH, Thelin A, Hansson L, Falk PG, Gordon JI. Molecular analysis of commensal host-microbial relationships in the intestine. Science 2001;291(5505):881–4. 链接1
[18] Smith PA. The tantalizing links between gut microbes and the brain. Nature 2015;526(7573):312–4. 链接1
[19] Tremaroli V, Bäckhed F. Functional interactions between the gut microbiota and host metabolism. Nature 2012;489(7415):242–9. 链接1
[20] Round JL, Mazmanian SK. The gut microbiota shapes intestinal immune responses during health and disease. Nat Rev Immunol 2009;9(5):313–23. 链接1
[21] Mazmanian SK, Liu CH, Tzianabos AO, Kasper DL. An immunomodulatory molecule of symbiotic bacteria directs maturation of the host immune system. Cell 2005;122(1):107–18. 链接1
[22] Bouskra D, Brézillon C, Bérard M, Werts C, Varona R, Boneca IG, et al. Lymphoid tissue genesis induced by commensals through NOD1 regulates intestinal homeostasis. Nature 2008;456(7221):507–10. 链接1
[23] Viaud S, Saccheri F, Mignot G, Yamazaki T, Daillère R, Hannani D, et al. The intestinal microbiota modulates the anticancer immune effects of cyclophosphamide. Science 2013;342(6161):971–6. 链接1
[24] Zeuthen LH, Fink LN, Frokiaer H. Epithelial cells prime the immune response to an array of gut-derived commensals towards a tolerogenic phenotype through distinct actions of thymic stromal lymphopoietin and transforming growth factor-beta. Immunology 2008;123(2):197–208. 链接1
[25] Massacand JC, Kaiser P, Ernst B, Tardivel A, Bürki K, Schneider P, et al. Intestinal bacteria condition dendritic cells to promote IgA production. PLoS One 2008;3(7):e2588. 链接1
[26] Kamada N, Chen GY, Inohara N, Núñez G. Control of pathogens and pathobionts by the gut microbiota. Nat Immunol 2013;14(7):685–90. 链接1
[27] Franks I. Microbiota: gut microbes might promote intestinal angiogenesis. Nat Rev Gastroenterol Hepatol 2013;10(1):3. 链接1
[28] Sharma R, Young C, Neu J. Molecular modulation of intestinal epithelial barrier: contribution of microbiota. J Biomed Biotechnol 2010;2010:305879. 链接1
[29] Freitas M, Axelsson LG, Cayuela C, Midtvedt T, Trugnan G. Microbial-host interactions specifically control the glycosylation pattern in intestinal mouse mucosa. Histochem Cell Biol 2002;118(2):149–61. 链接1
[30] Cryan JF, Dinan TG. Mind-altering microorganisms: the impact of the gut microbiota on brain and behaviour. Nat Rev Neurosci 2012;13 (10):701–12. 链接1
[31] Desbonnet L, Garrett L, Clarke G, Bienenstock J, Dinan TG. The probiotic Bifidobacteria infantis: an assessment of potential antidepressant properties in the rat. J Psychiatr Res 2008;43(2):164–74. 链接1
[32] Thomas RH, Meeking MM, Mepham JR, Tichenoff L, Possmayer F, Liu S, et al. The enteric bacterial metabolite propionic acid alters brain and plasma phospholipid molecular species: further development of a rodent model of autism spectrum disorders. J Neuroinflammation 2012;9(1):153. 链接1
[33] Qiu J. ‘Back to the future’ for Chinese herbal medicines. Nat Rev Drug Discov 2007;6(7):506–7. 链接1
[34] Wang HY, Qi LW, Wang CZ, Li P. Bioactivity enhancement of herbal supplements by intestinal microbiota focusing on ginsenosides. Am J Chin Med 2011;39(6):1103–15. 链接1
[35] Zhang X, Zhao Y, Zhang M, Pang X, Xu J, Kang C, et al. Structural changes of gut microbiota during berberine-mediated prevention of obesity and insulin resistance in high-fat diet-fed rats. PLoS ONE 2012;7(8):e42529. 链接1
[36] Cotillard A, Kennedy SP, Kong LC, Prifti E, Pons N, Le Chatelier E, et al. MicroObes consortium. Dietary intervention impact on gut microbial gene richness. Nature 2013;500(7464):585–8. 链接1
[37] Guo M, Ding S, Zhao C, Gu X, He X, Huang K, et al. Red ginseng and Semen Coicis can improve the structure of gut microbiota and relieve the symptoms of ulcerative colitis. J Ethnopharmacol 2015;162:7–13. 链接1
[38] Sekirov I, Russell SL, Antunes LC, Finlay BB. Gut microbiota in health and disease. Physiol Rev 2010;90(3):859–904. 链接1
[39] Marchesi JR, Adams DH, Fava F, Hermes GD, Hirschfield GM, Hold G, et al. The gut microbiota and host health: a new clinical frontier. Gut 2016;65 (2):330–9. 链接1
[40] Qin N, Yang F, Li A, Prifti E, Chen Y, Shao L, et al. Alterations of the human gut microbiome in liver cirrhosis. Nature 2014;513(7516):59–64. 链接1
[41] Chen Y, Yang F, Lu H, Wang B, Chen Y, Lei D, et al. Characterization of fecal microbial communities in patients with liver cirrhosis. Hepatology 2011;54 (2):562–72. 链接1
[42] Lu H, Wu Z, Xu W, Yang J, Chen Y, Li L. Intestinal microbiota was assessed in cirrhotic patients with hepatitis B virus infection. Intestinal microbiota of HBV cirrhotic patients. Microb Ecol 2011;61(3):693–703. 链接1
[43] Michail S, Lin M, Frey MR, Fanter R, Paliy O, Hilbush B, et al. Altered gut microbial energy and metabolism in children with non-alcoholic fatty liver disease. FEMS Microbiol Ecol 2015;91(2):1–9. 链接1
[44] Tang R, Wei Y, Li Y, Chen W, Chen H, Wang Q, et al. Gut microbial profile is altered in primary biliary cholangitis and partially restored after UDCA therapy. Gut 2018;67(3):534–41. 链接1
[45] Cao Y, Pan Q, Cai W, Shen F, Chen GY, Xu LM, et al. Modulation of gut microbiota by berberine improves steatohepatitis in high-fat diet-fed BALB/C mice. Arch Iran Med 2016;19(3):197–203. 链接1
[46] Yin X, Peng J, Zhao L, Yu Y, Zhang X, Liu P, et al. Structural changes of gut microbiota in a rat non-alcoholic fatty liver disease model treated with a Chinese herbal formula. Syst Appl Microbiol 2013;36(3):188–96. 链接1
[47] Dong H, Lu FE, Zhao L. Chinese herbal medicine in the treatment of nonalcoholic fatty liver disease. Chin J Integr Med 2012;18(2):152–60. 链接1
[48] Sakaida I, Tsuchiya M, Kawaguchi K, Kimura T, Terai S, Okita K. Herbal medicine Inchin-ko-to (TJ-135) prevents liver fibrosis and enzyme-altered lesions in rat liver cirrhosis induced by a choline-deficient L-amino aciddefined diet. J Hepatol 2003;38(6):762–9. 链接1
[49] Yamamoto M, Miura N, Ohtake N, Amagaya S, Ishige A, Sasaki H, et al. Genipin, a metabolite derived from the herbal medicine Inchin-ko-to, and suppression of Fas-induced lethal liver apoptosis in mice. Gastroenterology 2000;118(2):380–9. 链接1
[50] Yamamoto M, Ogawa K, Morita M, Fukuda K, Komatsu Y. The herbal medicine Inchin-ko-to inhibits liver cell apoptosis induced by transforming growth factor beta 1. Hepatology 1996;23(3):552–9. 链接1
[51] Yamashiki M, Mase A, Arai I, Huang XX, Nobori T, Nishimura A, et al. Effects of the Japanese herbal medicine ‘Inchinko-to’ (TJ-135) on concanavalin Ainduced hepatitis in mice. Clin Sci 2000;99(5):421–31. 链接1
[52] Huang W, Zhang J, Moore DD. A traditional herbal medicine enhances bilirubin clearance by activating the nuclear receptor CAR. J Clin Invest 2004;113(1):137–43. 链接1
[53] Liu P, Hu YY, Liu C, Zhu DY, Xue HM, Xu ZQ, et al. Clinical observation of salvianolic acid B in treatment of liver fibrosis in chronic hepatitis B. World J Gastroenterol 2002;8(4):679–85. 链接1
[54] Katiyar SK. Silymarin and skin cancer prevention: anti-inflammatory, antioxidant and immunomodulatory effects (Review). Int J Oncol 2005;26 (1):169–76. 链接1
[55] Lee JS, Kim SG, Kim HK, Lee TH, Jeong YI, Lee CM, et al. Silibinin polarizes Th1/ Th2 immune responses through the inhibition of immunostimulatory function of dendritic cells. J Cell Physiol 2007;210(2):385–97. 链接1
[56] Krecman V, Skottová N, Walterová D, Ulrichová J, Simánek V. Silymarin inhibits the development of diet-induced hypercholesterolemia in rats. Planta Med 1998;64(2):138–42. 链接1
[57] Skottová N, Krecman V. Silymarin as a potential hypocholesterolaemic drug. Physiol Res 1998;47(1):1–7. 链接1
[58] Wang S, Li X, Niu Y, Liu Y, Zhu Y, Lu X, et al. Identification and screening of chemical constituents with hepatoprotective effects from three traditional Chinese medicines for treating jaundice. J Sep Sci 2016;39(19):3690–9. 链接1
[59] Li Z, Yang S, Lin H, Huang J, Watkins PA, Moser AB, et al. Probiotics and antibodies to TNF inhibit inflammatory activity and improve nonalcoholic fatty liver disease. Hepatology 2003;37(2):343–50. 链接1
[60] Hsieh CY, Osaka T, Moriyama E, Date Y, Kikuchi J, Tsuneda S. Strengthening of the intestinal epithelial tight junction by Bifidobacterium bifidum. Physiol Rep 2015;3(3):e12327. 链接1
[61] Inao M, Mochida S, Matsui A, Eguchi Y, Yulutuz Y, Wang Y, et al. Japanese herbal medicine Inchin-ko-to as a therapeutic drug for liver fibrosis. J Hepatol 2004;41(4):584–91. 链接1
[62] Hackett ES, Twedt DC, Gustafson DL. Milk thistle and its derivative compounds: a review of opportunities for treatment of liver disease. J Vet Intern Med 2013;27(1):10–6. 链接1
[63] Fehér J, Láng I, Nékám K, Csomós G, Müzes G, Deák G. Effect of silibinin on the activity and expression of superoxide dismutase in lymphocytes from patients with chronic alcoholic liver disease. Free Radic Res Commun 1987;3(6):373–7. 链接1
[64] Zhang W, Hong R, Tian T. Silymarin’s protective effects and possible mechanisms on alcoholic fatty liver for rats. Biomol Ther 2013;21(4):264–9. 链接1
[65] Prakash P, Singh V, Jain M, Rana M, Khanna V, Barthwal MK, et al. Silymarin ameliorates fructose induced insulin resistance syndrome by reducing de novo hepatic lipogenesis in the rat. Eur J Pharmacol 2014;727:15–28. 链接1
[66] Feldstein AE, Canbay A, Angulo P, Taniai M, Burgart LJ, Lindor KD, et al. Hepatocyte apoptosis and fas expression are prominent features of human nonalcoholic steatohepatitis. Gastroenterology 2003;125(2):437–43. 链接1
[67] Akiho H, Yokoyama A, Abe S, Nakazono Y, Murakami M, Otsuka Y, et al. Promising biological therapies for ulcerative colitis: a review of the literature. World J Gastrointest Pathophysiol 2015;6(4):219–27. 链接1
[68] Ghouri YA, Richards DM, Rahimi EF, Krill JT, Jelinek KA, DuPont AW. Systematic review of randomized controlled trials of probiotics, prebiotics, and synbiotics in inflammatory bowel disease. Clin Exp Gastroenterol 2014;7:473–87. 链接1
[69] Nunes S, Danesi F, Del Rio D, Silva P. Resveratrol and inflammatory bowel disease: the evidence so far. Nutr Res Rev 2018;31(1):85–97. 链接1
[70] Zhang Z, Wu X, Cao S, Wang L, Wang D, Yang H, et al. Caffeic acid ameliorates colitis in association with increased Akkermansia population in the gut microbiota of mice. Oncotarget 2016;7(22):31790–9. 链接1
[71] Wang XM, Lu Y, Wu LY, Yu SG, Zhao BX, Hu HY, et al. Moxibustion inhibits interleukin-12 and tumor necrosis factor alpha and modulates intestinal flora in rat with ulcerative colitis. World J Gastroenterol 2012;18(46):6819–28. 链接1
[72] Wen J, Teng B, Yang P, Chen X, Li C, Jing Y, et al. The potential mechanism of Bawei Xileisan in the treatment of dextran sulfate sodium-induced ulcerative colitis in mice. J Ethnopharmacol 2016;188:31–8. 链接1
[73] Rahman H, Kim M, Leung G, Green JA, Katz S. Drug-herb interactions in the elderly patient with IBD: a growing concern. Curr Treat Options Gastroenterol 2017;15(4):618–36. 链接1
[74] Marchesi JR, Holmes E, Khan F, Kochhar S, Scanlan P, Shanahan F, et al. Rapid and noninvasive metabonomic characterization of inflammatory bowel disease. J Proteome Res 2007;6(2):546–51. 链接1
[75] Larrosa M, González-Sarrías A, Yáñez-Gascón MJ, Selma MV, Azorín-Ortuño M, Toti S, et al. Anti-inflammatory properties of a pomegranate extract and its metabolite urolithin-A in a colitis rat model and the effect of colon inflammation on phenolic metabolism. J Nutr Biochem 2010;21(8):717–25. 链接1
[76] Tchernof A, Després JP. Pathophysiology of human visceral obesity: an update. Physiol Rev 2013;93(1):359–404. 链接1
[77] Zhao L. The gut microbiota and obesity: from correlation to causality. Nat Rev Microbiol 2013;11(9):639–47. 链接1
[78] Itoh H. Gut microbiota and diseases: gut microbiota and obesity. Nippon Naika Gakkai Zasshi 2016;105(9):1712–6. 链接1
[79] Tang J, Feng Y, Tsao S, Wang N, Curtain R, Wang Y. Berberine and Coptidis rhizoma as novel antineoplastic agents: a review of traditional use and biomedical investigations. J Ethnopharmacol 2009;126(1):5–17. 链接1
[80] Li M, Shu X, Xu H, Zhang C, Yang L, Zhang L, et al. Integrative analysis of metabolome and gut microbiota in diet-induced hyperlipidemic rats treated with berberine compounds. J Transl Med 2016;14(1):237. 链接1
[81] Zhang X, Zhao Y, Xu J, Xue Z, Zhang M, Pang X, et al. Modulation of gut microbiota by berberine and metformin during the treatment of high-fat diet-induced obesity in rats. Sci Rep 2015;5(1):14405. 链接1
[82] Li X, Guo J, Ji K, Zhang P. Bamboo shoot fiber prevents obesity in mice by modulating the gut microbiota. Sci Rep 2016;6(1):32953. 链接1
[83] Bai J, Zhu Y, Dong Y. Modulation of gut microbiota and gut-generated metabolites by bitter melon results in improvement in the metabolic status in high fat diet-induced obese rats. J Funct Foods 2018;41:127–34. 链接1
[84] Sánchez-Tapia M, Aguilar-López M, Pérez-Cruz C, Pichardo-Ontiveros E, Wang M, Donovan SM, et al. Nopal (Opuntia ficus indica) protects from metabolic endotoxemia by modifying gut microbiota in obese rats fed high fat/sucrose diet. Sci Rep 2017;7(1):4716. 链接1
[85] Eid HM, Wright ML, Anil Kumar NV, Qawasmeh A, Hassan STS, Mocan A, et al. Significance of microbiota in obesity and metabolic diseases and the modulatory potential by medicinal plant and food ingredients. Front Pharmacol 2017;8:387. 链接1
[86] Han J, Lin H, Huang W. Modulating gut microbiota as an anti-diabetic mechanism of berberine. Med Sci Monit 2011;17(7):RA164–7. 链接1
[87] Xie W, Gu D, Li J, Cui K, Zhang Y. Effects and action mechanisms of berberine and Rhizoma coptidis on gut microbes and obesity in high-fat diet-fed C57BL/ 6J mice. PLoS ONE 2011;6(9):e24520. 链接1
[88] Crow JM. Microbiome: that healthy gut feeling. Nature 2011;480(7378): S88–9. 链接1
[89] Maslowski KM, Vieira AT, Ng A, Kranich J, Sierro F, Yu D, et al. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature 2009;461(7268):1282–6. 链接1
[90] Zhang Q, Piao XL, Piao XS, Lu T, Wang D, Kim SW. Preventive effect of Coptis chinensis and berberine on intestinal injury in rats challenged with lipopolysaccharides. Food Chem Toxicol 2011;49(1):61–9. 链接1
[91] Gu L, Li N, Gong J, Li Q, Zhu W, Li J. Berberine ameliorates intestinal epithelial tight-junction damage and down-regulates myosin light chain kinase pathways in a mouse model of endotoxinemia. J Infect Dis 2011;203 (11):1602–12. 链接1
[92] De Filippo C, Cavalieri D, Di Paola M, Ramazzotti M, Poullet JB, Massart S, et al. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc Natl Acad Sci USA 2010;107 (33):14691–6. 链接1
[93] Duncan SH, Lobley GE, Holtrop G, Ince J, Johnstone AM, Louis P, et al. Human colonic microbiota associated with diet, obesity and weight loss. Int J Obes 2008;32(11):1720–4. 链接1
[94] Li AN, Li S, Zhang YJ, Xu XR, Chen YM, Li HB. Resources and biological activities of natural polyphenols. Nutrients 2014;6(12):6020–47. 链接1
[95] Hanhineva K, Törrönen R, Bondia-Pons I, Pekkinen J, Kolehmainen M, Mykkänen H, et al. Impact of dietary polyphenols on carbohydrate metabolism. Int J Mol Sci 2010;11(4):1365–402. 链接1
[96] Cardona F, Andrés-Lacueva C, Tulipani S, Tinahones FJ, Queipo-Ortuño MI. Benefits of polyphenols on gut microbiota and implications in human health. J Nutr Biochem 2013;24(8):1415–22. 链接1
[97] Duda-Chodak A, Tarko T, Satora P, Sroka P. Interaction of dietary compounds, especially polyphenols, with the intestinal microbiota: a review. Eur J Nutr 2015;54(3):325–41. 链接1
[98] Hidalgo M, Oruna-Concha MJ, Kolida S, Walton GE, Kallithraka S, Spencer JPE, et al. Metabolism of anthocyanins by human gut microflora and their influence on gut bacterial growth. J Agric Food Chem 2012;60 (15):3882–90. 链接1
[99] Stevens JF, Maier CS. The chemistry of gut microbial metabolism of polyphenols. Phytochem Rev 2016;15(3):425–44. 链接1
[100] Kumar S, Pandey AK. Chemistry and biological activities of flavonoids: an overview. Sci World J 2013;2013:162750. 链接1
[101] Kwon O, Eck P, Chen S, Corpe CP, Lee JH, Kruhlak M, et al. Inhibition of the intestinal glucose transporter GLUT2 by flavonoids. FASEB J 2007;21 (2):366–77. 链接1
[102] Wang S, Moustaid-Moussa N, Chen L, Mo H, Shastri A, Su R, et al. Novel insights of dietary polyphenols and obesity. J Nutr Biochem 2014;25(1):1–18. 链接1
[103] Nie Q, Hu J, Gao H, Fan L, Chen H, Nie S. Polysaccharide from Plantago asiatica L. attenuates hyperglycemia, hyperlipidemia and affects colon microbiota in type 2 diabetic rats. Food Hydrocoll 2019;86:34–42. 链接1
[104] Liu G, Bei J, Liang L, Yu G, Li L, Li Q. Stachyose improves inflammation through modulating gut microbiota of high-fat diet/streptozotocin-induced type 2 diabetes in rats. Mol Nutr Food Res 2018;62(6):e1700954. 链接1
[105] Sung MM, Kim TT, Denou E, Soltys CM, Hamza SM, Byrne NJ, et al. Improved glucose homeostasis in obese mice treated with resveratrol is associated with alterations in the gut microbiome. Diabetes 2017;66(2):418–25. 链接1
[106] Martel J, Ojcius DM, Chang CJ, Lin CS, Lu CC, Ko YF, et al. Anti-obesogenic and antidiabetic effects of plants and mushrooms. Nat Rev Endocrinol 2017;13 (3):149–60. 链接1
[107] Wang JH, Bose S, Kim GC, Hong SU, Kim JH, Kim JE, et al. Flos Lonicera ameliorates obesity and associated endotoxemia in rats through modulation of gut permeability and intestinal microbiota. PLoS ONE 2014;9(1):e86117. 链接1
[108] Fang W, Wei C, Dong Y, Tang X, Zu Y, Chen Q. The effect on gut microbiota structure of primarily diagnosed type 2 diabetes patients intervened by Sancai Lianmei Particle and acarbose: a randomized controlled trial. J Clin Trials 2016;6(3):270. 链接1
[109] Xu J, Lian F, Zhao L, Zhao Y, Chen X, Zhang X, et al. Structural modulation of gut microbiota during alleviation of type 2 diabetes with a Chinese herbal formula. ISME J 2015;9(3):552–62. 链接1
[110] Chen X, D’Souza R, Hong ST. The role of gut microbiota in the gut-brain axis: current challenges and perspectives. Protein Cell 2013;4(6):403–14. 链接1
[111] Critchley JA, Zhang Y, Suthisisang CC, Chan TY, Tomlinson B. Alternative therapies and medical science: designing clinical trials of alternative/complementary medicines–is evidence-based traditional Chinese medicine attainable? J Clin Pharmacol 2000;40(5):462–7. 链接1











 京公网安备 11010502051620号
京公网安备 11010502051620号




