
2020年 第6卷 第4期
《工程(英文)》 >> 2020年 第6卷 第4期 doi: 10.1016/j.eng.2020.02.007
阪崎克罗诺杆菌生物膜形成机制及其在食品工业中的控制策略研究
a Guangdong Institute of Microbiology & Guangdong Academy of Sciences & State Key Laboratory of Applied Microbiology Southern China & Guangdong Provincial Key Laboratory of Microbiology Culture Collection and Application & Guangdong Open Laboratory of Applied Microbiology, Guangzhou 510070, China
b School of Bioscience and Bioengineering, South China University of Technology, Guangzhou 510006, China
c foodmicrobe.com, Keyworth, NG12 5GY, UK
d Department of Food Science and Technology, Jinan University, Guangzhou 510632, China
下一篇 上一篇
摘要
阪崎克罗诺杆菌(Cronobacter sakazakii)是一种食源性条件致病菌,可引起坏死性肠炎、脑膜炎和婴儿败血症等危及生命的侵袭性疾病。受该菌污染的婴幼儿配方奶粉(powdered infant formula, PIF)的潜在风险已引起乳品生产商、监管机构和消费者的广泛关注。C. sakazakii 可以在不同食品生产环境和设备表面形成一种保护该菌免受恶劣环境伤害的细胞生长模式——生物膜,并成为其在食品中持续污染的重要来源。然而,细菌生物膜由于其耐药特性难以去除。传统的清洗和消毒程序不足以完全控制生物膜,并且可能会导致进一步的生物膜形成和扩散。因此,基于纳米技术的药物传递系统、物种间相互作用以及制备来自微生物、天然提取物和噬菌体的抗菌分子等控制生物膜的新技术应运而生。本文将重点阐述C. sakazakii 生物膜形成机制及其潜在的控制策略。
图片

图1

图2

图3

图4

图5
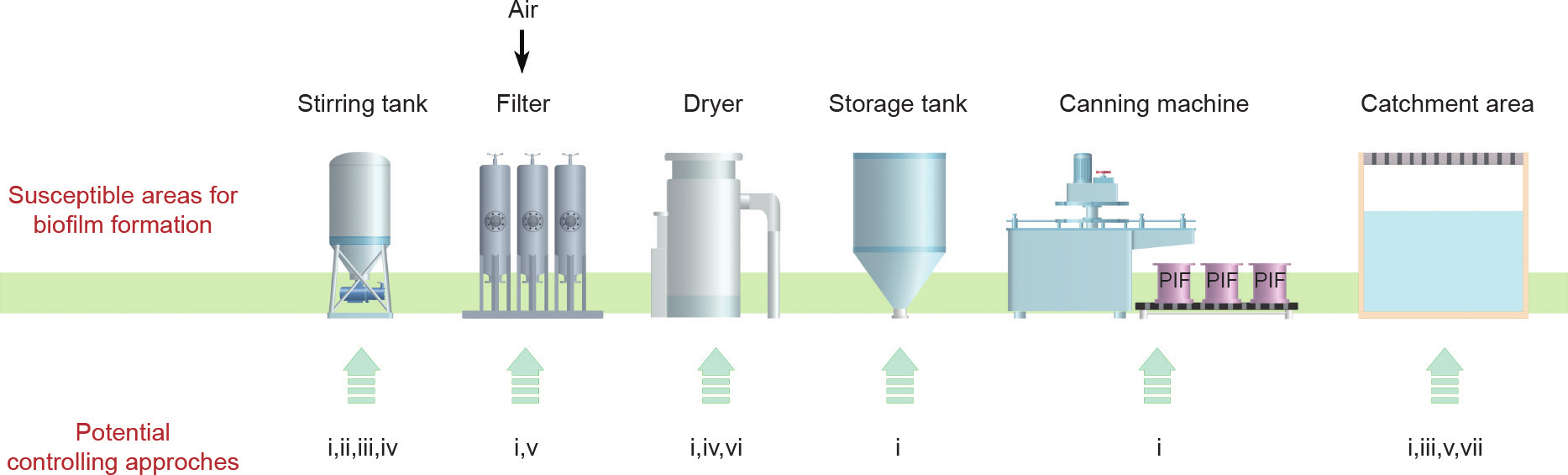
图6
参考文献
[ 1 ] Iversen C, Lehner A, Mullane N, Bidlas E, Cleenwerck I, Marugg J, et al. The taxonomy of Enterobacter sakazakii: proposal of a new genus Cronobacter gen. nov. and descriptions of Cronobacter sakazakii comb. nov. Cronobacter sakazakii subsp. sakazakii, comb. nov., Cronobacter sakazakii subsp. malonaticus subsp. nov., Cronobacter turicensis sp. nov., Cronobacter muytjensii sp. nov., Cronobacter dublinensis sp. nov. and Cronobacter genomospecies 1. BMC Evol Biol 2007;7(1):64. 链接1
[ 2 ] International Commission on Microbiological Specifications for Food. Microbiological testing in food safety management. Report. New York: Kluwer Academic/Plenum; 2002. 链接1
[ 3 ] Arseni A, Malamou-Ladas E, Koutsia C, Xanthou M, Trikka E. Outbreak of colonization of neonates with Enterobacter sakazakii. J Hosp Infect 1987;9 (2):143–50. 链接1
[ 4 ] Centers for Disease Control Prevention. Cronobacter species isolation in two infants—New Mexico, 2008. MMWR Morb Mortal Wkly Rep 2009;58:1179–83. 链接1
[ 5 ] Lai KK. Enterobacter sakazakii infections among neonates, infants, children, and adults. Case reports and a review of the literature. Medicine 2001;80 (2):113–22. 链接1
[ 6 ] Ries M, Harms D, Scharf J. Multiple cerebral infarcts with resulting multicystic encephalomalacia in a premature infant with Enterobacter sakazakii meningitis. Klin Padiatr 1994;206(3):184–6. 链接1
[ 7 ] Van Acker J, De Smet F, Muyldermans G, Bougatef A, Naessens A, Lauwers S. Outbreak of necrotizing enterocolitis associated with Enterobacter sakazakii in powdered milk formula. J Clin Microbiol 2001;39(1):293–7. 链接1
[ 8 ] Yong W, Guo B, Shi X, Cheng T, Chen M, Jiang X, et al. An investigation of an acute gastroenteritis outbreak: Cronobacter sakazakii, a potential cause of food-borne illness. Front Microbiol 2018;9:2549. 链接1
[ 9 ] Patrick ME, Mahon BE, Greene SA, Rounds J, Cronquist A, Wymore K, et al. Incidence of Cronobacter spp. infections, United States, 2003–2009. Emerg Infect Dis 2014;20(9):1520–3. 链接1
[10] Belal M, Al-Mariri A, Hallab L, Hamad I. Detection of Cronobacter spp. (formerly Enterobacter sakazakii) from medicinal plants and spices in Syria. J Infect Dev Ctries 2013;7(2):82–9. 链接1
[11] Fei P, Jiang Y, Gong S, Li R, Jiang Y, Yuan X, et al. Genotyping occurrence and antibiotic susceptibility of Cronobacter spp. in drinking water and food samples from Northeast, China. J Food Prot 2018;81(3):456–60. 链接1
[12] Forsythe SJ. Enterobacter sakazakii and other bacteria in powdered infant milk formula. Matern Child Nutr 2005;1(1):44–50. 链接1
[13] Ling N, Li C, Zhang J, Wu Q, Zeng H, He W, et al. Prevalence and molecular and antimicrobial characteristics of Cronobacter spp. isolated from raw vegetables in China. Front Microbiol 2018;9:1149. 链接1
[14] Ueda S. Occurrence of Cronobacter spp. in dried foods, fresh vegetables and soil. Biocontrol Sci 2017;22(1):55–9. 链接1
[15] Vasconcellos L, Carvalho CT, Tavares RO, De Mello Medeiros V, De Oliveira Rosas C, Silva JN, et al. Isolation, molecular and phenotypic characterization of Cronobacter spp. in ready-to-eat salads and foods from Japanese cuisine commercialized in Brazil. Food Res Int 2018;107:353–9. 链接1
[16] Xu X, Wu Q, Zhang J, Ye Y, Yang X, Dong X. Occurrence and characterization of Cronobacter spp. in powdered formula from Chinese retail markets. Foodborne Pathog Dis 2014;11(4):307–12. 链接1
[17] Ye Y, Li H, Wu Q, Chen M, Lu Y, Yan C. Isolation and phenotypic characterization of Cronobacter from dried edible macrofungi samples. J Food Sci 2014;79(7):M1382–6. 链接1
[18] Hamilton JV, Lehane MJ, Braig HR. Isolation of Enterobacter sakazakii from midgut of Stomoxys calcitrans. Emerg Infect Dis 2003;9(10):1355–6. 链接1
[19] Pava-Ripoll M, Pearson RE, Miller AK, Ziobro GC. Prevalence and relative risk of Cronobacter spp., Salmonella spp., and Listeria monocytogenes associated with the body surfaces and guts of individual filth flies. Appl Environ Microbiol 2012;78(22):7891–902. 链接1
[20] Iversen C, Forsythe S. Risk profile of Enterobacter sakazakii, an emergent pathogen associated with infant milk formula. Trends Food Sci Technol 2003;14(11):443–54. 链接1
[21] Kandhai MC, Reij MW, Gorris LG, Guillaume-Gentil O, Van Schothorst M. Occurrence of Enterobacter sakazakii in food production environments and households. Lancet 2004;363(9402):39–40. 链接1
[22] Killer J, Skrˇivanová E, Hochel I, Marounek M. Multilocus sequence typing of Cronobacter strains isolated from retail foods and environmental samples. Foodborne Pathog Dis 2015;12(6):514–21. 链接1
[23] Arts M. Enterobacter sakazakii in factories and households. Lancet 2004;364 (9432):414–5. 链接1
[24] Fang R, Wang Q, Yang B, Zhang J, Cao B, Geng W, et al. Prevalence and subtyping of Cronobacter species in goat milk powder factories in Shaanxi Province, China. J Dairy Sci 2015;98(11):7552–9. 链接1
[25] Hall-Stoodley L, Costerton JW, Stoodley P. Bacterial biofilms: from the natural environment to infectious diseases. Nat Rev Microbiol 2004;2 (2):95–108. 链接1
[26] Prosser JI, Bohannan BJM, Curtis TP, Ellis RJ, Firestone MK, Freckleton RP, et al. The role of ecological theory in microbial ecology. Nat Rev Microbiol 2007;5 (5):384–92. 链接1
[27] Boles BR, Thoendel M, Singh PK. Self-generated diversity produces ‘‘insurance effects” in biofilm communities. Proc Natl Acad Sci USA 2004;101(47):16630–5. 链接1
[28] Flemming HC, Wingender J. The biofilm matrix. Nat Rev Microbiol 2010;8 (9):623–33. 链接1
[29] Iversen C, Lane M, Forsythe SJ. The growth profile, thermotolerance and biofilm formation of Enterobacter sakazakii grown in infant formula milk. Lett Appl Microbiol 2004;38(5):378–82. 链接1
[30] Lehner A, Riedel K, Eberl L, Breeuwer P, Diep B, Stephan R. Biofilm formation, extracellular polysaccharide production, and cell-to-cell signaling in various Enterobacter sakazakii strains: aspects promoting environmental persistence. J Food Prot 2005;68(11):2287–94. 链接1
[31] Beuchat LR, Kim H, Gurtler JB, Lin LC, Ryu JH, Richards GM. Cronobacter sakazakii in foods and factors affecting its survival, growth, and inactivation. Int J Food Microbiol 2009;136(2):204–13. 链接1
[32] Hurrell E, Kucerova E, Loughlin M, Caubilla-Barron J, Forsythe SJ. Biofilm formation on enteral feeding tubes by Cronobacter sakazakii, Salmonella serovars and other Enterobacteriaceae. Int J Food Microbiol 2009;136 (2):227–31. 链接1
[33] Kragh KN, Hutchison JB, Melaugh G, Rodesney C, Roberts AE, Irie Y, et al. Role of multicellular aggregates in biofilm formation. MBio 2016;7(2): e00237–316. 链接1
[34] Ling N, Zhang J, Li C, Zeng H, He W, Ye Y, et al. The glutaredoxin gene, grxB, affects acid tolerance, surface hydrophobicity, auto-aggregation, and biofilm formation in Cronobacter sakazakii. Front Microbiol 2018;9:133. 链接1
[35] Jung JH, Choi NY, Lee SY. Biofilm formation and exopolysaccharide (EPS) production by Cronobacter sakazakii depending on environmental conditions. Food Microbiol 2013;34(1):70–80. 链接1
[36] Ye Y, Ling N, Jiao R, Wu Q, Han Y, Gao J. Effects of culture conditions on the biofilm formation of Cronobacter sakazakii strains and distribution of genes involved in biofilm formation. Lebensm Wiss Technol 2015;62(1):1–6. 链接1
[37] Dancer GI, Mah JH, Kang DH. Influences of milk components on biofilm formation of Cronobacter spp. (Enterobacter sakazakii). Lett Appl Microbiol 2009;48(6):718–25. 链接1
[38] Ye Y, Ling N, Jiao R, Wu Q, Han Y, Gao J. Effects of Ca2+ and Mg2+ on the biofilm formation of Cronobacter sakazakii strains from powdered infant formula. J Food Saf 2015;35(3):416–21. 链接1
[39] Soni KA, Balasubramanian AK, Beskok A, Pillai SD. Zeta potential of selected bacteria in drinking water when dead, starved, or exposed to minimal and rich culture media. Curr Microbiol 2008;56(1):93–7. 链接1
[40] Pringle JH, Fletcher M. Influence of substratum wettability on attachment of freshwater bacteria to solid surfaces. Appl Environ Microbiol 1983;45 (3):811–7. 链接1
[41] Thomas WE, Nilsson LM, Forero M, Sokurenko EV, Vogel V. Shear-dependent, ‘‘stick-and-roll” adhesion of type 1 fimbriated Escherichia coli. Mol Microbiol 2004;53(5):1545–57. 链接1
[42] Davidson CAB, Lowe CR. Optimisation of polymeric surface pre-treatment to prevent bacterial biofilm formation for use in microfluidics. J Mol Recognit 2004;17(3):180–5. 链接1
[43] Heistad A, Scott T, Skaarer AM, Seidu R, Hanssen JF, Stenström TA, et al. Virus removal by unsaturated wastewater filtration: effects of biofilm accumulation and hydrophobicity. Water Sci Technol 2009;60(2):399–407. 链接1
[44] Nadell CD, Xavier JB, Levin SA, Foster KR. The evolution of quorum sensing in bacterial biofilms. PLoS Biol 2008;6(1):e14. 链接1
[45] Teschler JK, Zamorano-Sánchez D, Utada AS, Warner CJ, Wong GC, Linington RG, et al. Living in the matrix: assembly and control of Vibrio cholerae biofilms. Nat Rev Microbiol 2015;13(5):255–68. 链接1
[46] Jayaraman A, Wood TK. Bacterial quorum sensing: signals, circuits, and implications for biofilms and disease. Annu Rev Biomed Eng 2008;10 (1):145–67. 链接1
[47] Fuqua C, Winans SC, Greenberg EP. Census and consensus in bacterial ecosystems: the LuxR-LuxI family of quorum-sensing transcriptional regulators. Annu Rev Microbiol 1996;50(1):727–51. 链接1
[48] Dickschat JS. Quorum sensing and bacterial biofilms. Nat Prod Rep 2010;27 (3):343–69. 链接1
[49] Da Silva Araújo FD, Esper LMR, Kuaye AY, Sircili MP. Marsaioli AJ. N-acylhomoserine lactones from Enterobacter sakazakii (Cronobacter spp.) and their degradation by Bacillus cereus enzymes. J Agric Food Chem 2012;60 (2):585–92. 链接1
[50] Singh N, Patil A, Prabhune A, Raghav M, Goel G. Diverse profiles of N-acylhomoserine lactones in biofilm forming strains of Cronobacter sakazakii. Virulence 2017;8(3):275–81. 链接1
[51] Du XJ, Wang F, Lu X, Rasco BA, Wang S. Biochemical and genetic characteristics of Cronobacter sakazakii biofilm formation. Res Microbiol 2012;163(6–7):448–56. 链接1
[52] Yang JL, He YP, Jiang J, Chen WY, Gao Q, Pan LW, et al. Comparative proteomic analysis by iTRAQ-2DLC-MS/MS provides insight into the key proteins involved in Cronobacter spp. biofilm formation. Food Control 2016;63:93–100. 链接1
[53] Ye YW, Jiao R, Gao JN, Li H, Ling N, Wu QP, et al. Proteins involved in responses to biofilm and planktonic modes in Cronobacter sakazakii. Lebensm Wiss Technol 2016;65:1093–9. 链接1
[54] Ogrodzki P, Forsythe S. Capsular profiling of the Cronobacter genus and the association of specific Cronobacter sakazakii and C. malonaticus capsule types with neonatal meningitis and necrotizing enterocolitis. BMC Genomics 2015;16(1):758. 链接1
[55] Hu L, Grim CJ, Franco AA, Jarvis KG, Sathyamoorthy V, Kothary MH, et al. Analysis of the cellulose synthase operon genes, bcsA, bcsB, and bcsC in Cronobacter species: prevalence among species and their roles in biofilm formation and cell–cell aggregation. Food Microbiol 2015;52:97–105. 链接1
[56] Gao JX, Li P, Du XJ, Han ZH, Xue R, Liang B, et al. A negative regulator of cellulose biosynthesis, bcsR, affects biofilm formation, and adhesion/invasion ability of Cronobacter sakazakii. Front Microbiol 2017;8:1839. 链接1
[57] Bullitt E, Makowski L. Structural polymorphism of bacterial adhesion pili. Nature 1995;373(6510):164–7. 链接1
[58] Joseph S, Desai P, Ji Y, Cummings CA, Shih R, Degoricija L, et al. Comparative analysis of genome sequences covering the seven cronobacter species. PLoS ONE 2012;7(11):e49455. 链接1
[59] DeBenedictis EP, Liu J, Keten S. Adhesion mechanisms of curli subunit CsgA to abiotic surfaces. Sci Adv 2016;2(11):e1600998. 链接1
[60] Hartmann I, Carranza P, Lehner A, Stephan R, Eberl L, Riedel K. Genes involved in Cronobacter sakazakii biofilm formation. Appl Environ Microbiol 2010;76 (7):2251–61. 链接1
[61] Chapman MR, Robinson LS, Pinkner JS, Roth R, Heuser J, Hammar M, et al. Role of Escherichia coli curli operons in directing amyloid fiber formation. Science 2002;295(5556):851–5. 链接1
[62] Hammar M, Arnqvist A, Bian Z, Olsén A, Normark S. Expression of two csg operons is required for production of fibronectin- and Congo redbinding curli polymers in Escherichia coli K-12. Mol Microbiol 1995;18 (4):661–70. 链接1
[63] Hu L. Prevalence of curli genes among Cronobacter species and their roles in biofilm formation and cell–cell aggregation. Int J Food Microbiol 2018; 265:65–73. 链接1
[64] Lemon KP, Higgins DE, Kolter R. Flagellar motility is critical for Listeria monocytogenes biofilm formation. J Bacteriol 2007;189(12):4418–24. 链接1
[65] Zhao X, Norris SJ, Liu J. Molecular architecture of the bacterial flagellar motor in cells. Biochemistry 2014;53(27):4323–33. 链接1
[66] Macnab RM. How bacteria assemble flagella. Annu Rev Microbiol 2003;57 (1):77–100. 链接1
[67] Hoeflinger JL, Miller MJ. Cronobacter sakazakii ATCC 29544 autoaggregation requires FliC flagellation, not motility. Front Microbiol 2017;8:301. 链接1
[68] Franzosa EA, Hsu T, Sirota-Madi A, Shafquat A, Abu-Ali G, Morgan XC, et al. Sequencing and beyond: integrating molecular ‘‘omics” for microbial community profiling. Nat Rev Microbiol 2015;13(6):360–72. 链接1
[69] Papenfort K, Förstner KU, Cong JP, Sharma CM, Bassler BL. Differential RNAseq of Vibrio cholerae identifies the VqmR small RNA as a regulator of biofilm formation. Proc Natl Acad Sci USA 2015;112(7):E766–75. 链接1
[70] Johnson CH, Ivanisevic J, Siuzdak G. Metabolomics: beyond biomarkers and towards mechanisms. Nat Rev Mol Cell Biol 2016;17(7):451–9. 链接1
[71] Fei P, Man C, Lou B, Forsythe SJ, Chai Y, Li R, et al. Genotyping and source tracking of the Cronobacter sakazakii and C. malonaticus isolated from powdered infant formula and an infant formula production factory in China. Appl Environ Micob 2015;81(16):5430–9. 链接1
[72] Jacobs C, Braun P, Hammer P. Reservoir and routes of transmission of Enterobacter sakazakii (Cronobacter spp.) in a milk powder-producing plant. J Dairy Sci 2011;94(8):3801–10. 链接1
[73] Reich F, König R. Von Wiese W, Klein G. Prevalence of Cronobacter spp. in a powdered infant formula processing environment. Int J Food Microbiol 2010;140(2–3):214–7. 链接1
[74] FAO. Code of hygienic practice for powdered formulae for infants and young children. Report. Geneva: Food and Agriculture Organization of the United Nation; 2008. Report No.: CAC/RCP 66-2008.
[75] Cordier JL. Production of powdered infant formulae and microbiological control measures. In: Farber JM, Forsythe SJ, editors. Emerging issues in food safety: Enterobacter sakazakii. Washington, DC: ASM Press; 2007. 链接1
[76] World Health Organization, Food and Agriculture Organization of the United Nations. Enterobacter sakazakii and other microorganisms in powdered infant formula. Geneva: World Health Organization; 2004.
[77] Arku B, Fanning S, Jordan K. Heat adaptation and survival of Cronobacter spp. (formerly Enterobacter sakazakii). Foodborne Pathog Dis 2011;8(9): 975–81. 链接1
[78] Kim H, Ryu JH, Beuchat LR. Effectiveness of disinfectants in killing Enterobacter sakazakii in suspension, dried on the surface of stainless steel, and in a biofilm. Appl Environ Microbiol 2007;73(4):1256–65. 链接1
[79] Thomas A, Sathian CT. Cleaning-in-place (CIP) system in dairy plant—review. J Environ Sci Toxicol Food Technol 2014;8(6):41–4. 链接1
[80] Bremer PJ, Fillery S, McQuillan AJ. Laboratory scale clean-in-place (CIP) studies on the effectiveness of different caustic and acid wash steps on the removal of dairy biofilms. Int J Food Microbiol 2006;106(3):254–62. 链接1
[81] Boulange-Petermann L, Jullien C, Dubois PE, Benezech T, Faille C. Influence of surface chemistry on the hygienic status of industrial stainless steel. Biofouling 2004;20(1):25–33. 链接1
[82] Lelievre C, Faille C, Benezech T. Removal kinetics of Bacillus cereus, spores from stainless steel pipes under CIP procedure: influence of soiling and cleaning conditions. J Food Process Eng 2001;24(6):359–79. 链接1
[83] Lelièvre C, Legentilhomme P, Gaucher C, Legrand J, Faille C, Bénézech T. Cleaning in place: effect of local wall shear stress variation on bacterial removal from stainless steel equipment. Chem Eng Sci 2002;57 (8):1287–97. 链接1
[84] Kumari S, Sarkar PK. In vitro model study for biofilm formation by Bacillus cereus in dairy chilling tanks and optimization of clean-in-place (CIP) regimes using response surface methodology. Food Control 2014;36 (1):153–8. 链接1
[85] Elguindi J, Moffitt S, Hasman H, Andrade C, Raghavan S, Rensing C. Metallic copper corrosion rates, moisture content, and growth medium influence survival of copper ion-resistant bacteria. Appl Microbiol Biotechnol 2011;89 (6):1963–70. 链接1
[86] Grass G, Rensing C, Solioz M. Metallic copper as an antimicrobial surface. Appl Environ Microbiol 2011;77(5):1541–7. 链接1
[87] Elguindi J, Alwathnani HA, Rensing C. Rapid inactivation of Cronobacter sakazakii on copper alloys following periods of desiccation stress. World J Microbiol Biotechnol 2012;28(4):1837–41. 链接1
[88] Edelson-Mammel SG, Porteous MK, Buchanan RL. Survival of Enterobacter sakazakii in a dehydrated powdered infant formula. J Food Prot 2005;68 (9):1900–2. 链接1
[89] Breeuwer P, Lardeau A, Peterz M, Joosten HM. Desiccation and heat tolerance of Enterobacter sakazakii. J Appl Microbiol 2003;95(5):967–73. 链接1
[90] Nazarowec-White M, Farber JM. Thermal resistance of Enterobacter sakazakii in reconstituted dried-infant formula. Lett Appl Microbiol 1997;24 (1):9–13. 链接1
[91] Iversen C, Druggan P, Forsythe S. A selective differential medium for Enterobacter sakazakii, a preliminary study. Int J Food Microbiol 2004;96 (2):133–9. 链接1
[92] Awadallah MAI, Ahmed HA, Merwad AMA, Abou Elez RMM, Saleh KMA. Molecular characterization of Cronobacter sakazakii in Egypt, survival and thermoresistance at different temperatures: a potential public health risk. Vector Borne Zoonotic Dis 2018;18(2):101–7. 链接1
[93] Ueda S. The effects of temperature on the growth and heat resistance of Cronobacter spp. Biocontrol Sci 2017;22(2):125–9. 链接1
[94] Arku B, Mullane N, Fox EM, Fanning S, Jordan KN. Enterobacter sakazakii survives spray drying. Int J Dairy Sci 2008;61(1):102–8. 链接1
[95] Mahmoud BS. Inactivation effect of X-ray treatments on Cronobacter species (Enterobacter sakazakii) in tryptic soy broth, skim milk, low-fat milk and whole-fat milk. Lett Appl Microbiol 2009;49(5):562–7. 链接1
[96] Santo D, Graça A, Nunes C, Quintas C. Survival and growth of Cronobacter sakazakii on fresh-cut fruit and the effect of UV-C illumination and electrolyzed water in the reduction of its population. Int J Food Microbiol 2016;231:10–5. 链接1
[97] Ha JW, Kang DH. Synergistic bactericidal effect of simultaneous near-infrared radiant heating and UV radiation against Cronobacter sakazakii in powdered infant formula. Appl Environ Microbiol 2014;80(6):1858–63. 链接1
[98] Oulahal-Lagsir N, Martial-Gros A, Bonneau M, Blum LJ. Ultrasonic methodology coupled to ATP bioluminescence for the non-invasive detection of fouling in food processing equipment—validation and application to a dairy factory. J Appl Microbiol 2000;89(3):433–41. 链接1
[99] Adekunte A, Valdramidis VP, Tiwari BK, Slone N, Cullen PJ, Donnell CP, et al. Resistance of Cronobacter sakazakii in reconstituted powdered infant formula during ultrasound at controlled temperatures: a quantitative approach on microbial responses. Int J Food Microbiol 2010;142(1–2):53–9. 链接1
[100] Arroyo C, Cebrián G, Pagán R, Condón S. Inactivation of Cronobacter sakazakii by ultrasonic waves under pressure in buffer and foods. Int J Food Microbiol 2011;144(3):446–54. 链接1
[101] Torlak E, Gokmen M, Aydemir S. Efficacy of disinfectants against Cronobacter biofilm on plastic surfaces. Qual Assur Saf Crop 2015;7 (5):819–23. 链接1
[102] Condell O, Iversen C, Cooney S, Power KA, Walsh C, Burgess C, et al. Efficacy of biocides used in the modern food industry to control salmonella enterica, and links between biocide tolerance and resistance to clinically relevant antimicrobial compounds. Appl Environ Microbiol 2012;78(9): 3087–3097. 链接1
[103] Amalaradjou MAR, Venkitanarayanan K. Effect of trans-cinnamaldehyde on inhibition and inactivation of Cronobacter sakazakii biofilm on abiotic surfaces. J Food Prot 2011;74(2):200–8. 链接1
[104] Amalaradjou MA, Kim KS, Venkitanarayanan K. Sub-inhibitory concentrations of trans-cinnamaldehyde attenuate virulence in Cronobacter sakazakii in vitro. Int J Mol Sci 2014;15(5):8639–55. 链接1
[105] Kim TJ, Silva JL, Weng WL, Chen WW, Corbitt M, Jung YS, et al. Inactivation of Enterobacter sakazakii by water-soluble muscadine seed extracts. Int J Food Microbiol 2009;129(3):295–9. 链接1
[106] Joshi SS, Howell AB, D’Souza DH. Cronobacter sakazakii reduction by blueberry proanthocyanidins. Food Microbiol 2014;39:127–31. 链接1
[107] Hancock REW, Sahl HG. Antimicrobial and host-defense peptides as new antiinfective therapeutic strategies. Nat Biotechnol 2006;24(12):1551–7. 链接1
[108] Melo MN, Ferre R, Castanho MARB. Antimicrobial peptides: linking partition, activity and high membrane-bound concentrations. Nat Rev Microbiol 2009;7(3):245–50. 链接1
[109] Rai M, Pandit R, Gaikwad S, Kövics G. Antimicrobial peptides as natural biopreservative to enhance the shelf-life of food. J Food Sci Technol 2016;53 (9):3381–94. 链接1
[110] Upendra RS, Khandelwal P, Jana K, Ajay Kumar N, Gayathri Devi M, Stephaney ML. Bacteriocin production from indigenous strains of lactic acid bacteria isolated from selected fermented food sources. Int J Pharma Res Health Sci 2016;4(1):982–90. 链接1
[111] Jorge P, Lourenço A, Pereira MO. New trends in peptide-based anti-biofilm strategies: a review of recent achievements and bioinformatic approaches. Biofouling 2012;28(10):1033–61. 链接1
[112] Di Luca M, Maccari G, Nifosì R. Treatment of microbial biofilms in the postantibiotic era: prophylactic and therapeutic use of antimicrobial peptides and their design by bioinformatics tools. Pathog Dis 2014;70(3):257–70. 链接1
[113] Hell Ã, Giske CG, Nelson A, Römling U, Marchini G. Human cathelicidin peptide LL37 inhibits both attachment capability and biofilm formation of Staphylococcus epidermidis. Lett Appl Microbiol 2010;50(2):211–5. 链接1
[114] Overhage J, Campisano A, Bains M, Torfs EC, Rehm BH, Hancock RE. Human host defense peptide LL-37 prevents bacterial biofilm formation. Infect Immun 2008;76(9):4176–82. 链接1
[115] Kim DH, Jeong D, Song KY, Kang IB, Kim H, Seo KH. Culture supernatant produced by Lactobacillus kefiri from kefir inhibits the growth of Cronobacter sakazakii. J Dairy Res 2018;85(1):98–103. 链接1
[116] Awaisheh SS, Al-Nabulsi AA, Osaili TM, Ibrahim S, Holley R. Inhibition of Cronobacter sakazakii by heat labile bacteriocins produced by probiotic LAB isolated from healthy infants. J Food Sci 2013;78(9):M1416–20. 链接1
[117] Kudva IT, Jelacic S, Tarr PI, Youderian P, Hovde CJ. Biocontrol of Escherichia coli O157 with O157-specific bacteriophages. Appl Environ Microbiol 1999;65(9):3767–73. 链接1
[118] Rossmann FS, Racek T, Wobser D, Puchalka J, Rabener EM, Reiger M, et al. Phage-mediated dispersal of biofilm and distribution of bacterial virulence genes is induced by quorum sensing. PLoS Pathog 2015;11(2):e1004653. 链接1
[119] Bren L. Bacteria-eating virus approved as food additive. FDA Consum 2007;41 (1):20–2. 链接1
[120] Breeuwer P, Boissindelaporte C, Joosten H, Lardeau A. Isolated phages and their use as disinfectant in food or for sanitation of factory environment. Switzerland patent WO2005049813A1P. 2003 Nov 19.
[121] Endersen L, Buttimer C, Nevin E, Coffey A, Neve H, Oliveira H, et al. Investigating the biocontrol and anti-biofilm potential of a three phage cocktail against Cronobacter sakazakii in different brands of infant formula. Int J Food Microbiol 2017;253:1–11. 链接1
[122] Rizzello L, Cingolani R, Pompa PP. Nanotechnology tools for antibacterial materials. Nanomedicine 2013;8(5):807–21. 链接1
[123] Besinis A, Hadi SD, Le HR, Tredwin C, Handy RD. Antibacterial activity and biofilm inhibition by surface modified titanium alloy medical implants following application of silver, titanium dioxide and hydroxyapatite nanocoatings. Nanotoxicology 2017;11(3):327–38. 链接1
[124] Pelgrift RY, Friedman AJ. Nanotechnology as a therapeutic tool to combat microbial resistance. Adv Drug Deliv Rev 2013;65(13–14): 1803–15. 链接1
[125] Dos Santos Ramos MA, Da Silva PB, Spósito L, De Toledo LG, Bonifácio BV, Rodero CF, et al. Nanotechnology-based drug delivery systems for control of microbial biofilms: a review. Int J Nanomed 2018;13:1179–213. 链接1
[126] Robinson AM, Bannister M, Creeth JE, Jones MN. The interaction of phospholipid liposomes with mixed bacterial biofilms and their use in the delivery of bactericide. Colloid Surface A 2001;186(1–2):43–53. 链接1
[127] Wei H, Wang E. Nanomaterials with enzyme-like characteristics (nanozymes): next-generation artificial enzymes. Chem Soc Rev 2013;42 (14):6060–93. 链接1
[128] Tao Y, Ju E, Ren J, Qu X. Bifunctionalized mesoporous silica-supported gold nanoparticles: intrinsic oxidase and peroxidase catalytic activities for antibacterial applications. Adv Mater 2015;27(6):1097–104. 链接1
[129] Gao L, Giglio KM, Nelson JL, Sondermann H, Travis AJ. Ferromagnetic nanoparticles with peroxidase-like activity enhance the cleavage of biological macromolecules for biofilm elimination. Nanoscale 2014;6 (5):2588–93. 链接1
[130] Amara N, Krom BP, Kaufmann GF, Meijler MM. Macromolecular inhibition of quorum sensing: enzymes, antibodies, and beyond. Chem Rev 2011;111 (1):195–208. 链接1
[131] Singh N, Patil A, Prabhune A, Goel G. Inhibition of quorum-sensing-mediated biofilm formation in Cronobacter sakazakii strains. Microbiology 2016;162 (9):1708–14. 链接1











 京公网安备 11010502051620号
京公网安备 11010502051620号




