
2022, Volume 17, Issue 10
Engineering >> 2022, Volume 17, Issue 10 doi: 10.1016/j.eng.2021.05.009
Identification of Monocytes Associated with Severe COVID-19 in the PBMCs of Severely Infected Patients Through Single-Cell Transcriptome Sequencing
a State Key Laboratory for Diagnosis and Treatment of Infectious Disease, National Clinical Research Center for Infectious Diseases, Collaborative Innovation Center for Diagnosis and Treatment of Infectious Diseases, Department of Infectious Disease, The First Affiliated Hospital, College of Medicine, Zhejiang University, Hangzhou 310003, China
b Shulan (Hangzhou) Hospital Affiliated to Zhejiang Shuren University Shulan International Medical College, Hangzhou 310022, China
# These authors contributed equally to this work.
Next Previous
Abstract
Understanding the immunological characteristics of monocytes—including the characteristics associated with fibrosis—in severe coronavirus disease 2019 (COVID-19) is crucial for understanding the pathogenic mechanism of the disease and preventing disease severity. In this study, we performed single-cell transcriptomic sequencing of peripheral blood samples collected from six healthy controls and 14 COVID-19 samples including severe, moderate, and convalescent samples from three severely/critically ill and four moderately ill patients. We found that the monocytes were strongly remodeled in the severely/critically ill patients with COVID-19, with an increased proportion of monocytes and seriously reduced diversity. In addition, we discovered two novel severe-disease-specific monocyte subsets: Mono 0 and Mono 5. These subsets expressed amphiregulin (AREG), epiregulin (EREG), and cytokine interleukin-18 (IL-18) gene, exhibited an enriched erythroblastic leukemia viral oncogene homolog (ErbB) signaling pathway, and appeared to exhibit pro-fibrogenic and pro-inflammation characteristics. We also found metabolic changes in Mono 0 and Mono 5, including increased glycolysis/gluconeogenesis and an increased hypoxia inducible factor-1 (HIF-1) signaling pathway. Notably, one pre-severe sample displayed a monocyte atlas similar to that of the severe/critical samples. In conclusion, our study discovered two novel severe-disease-specific monocyte subsets as potential predictors and therapeutic targets for severe COVID-19. Overall, this study provides potential predictors for severe disease and therapeutic targets for COVID-19 and thus provides a resource for further studies on COVID-19.
Keywords
COVID-19 ; Severe infection ; Critically ill ; Monocytes ; Fibrosis
SupplementaryMaterials
Figures
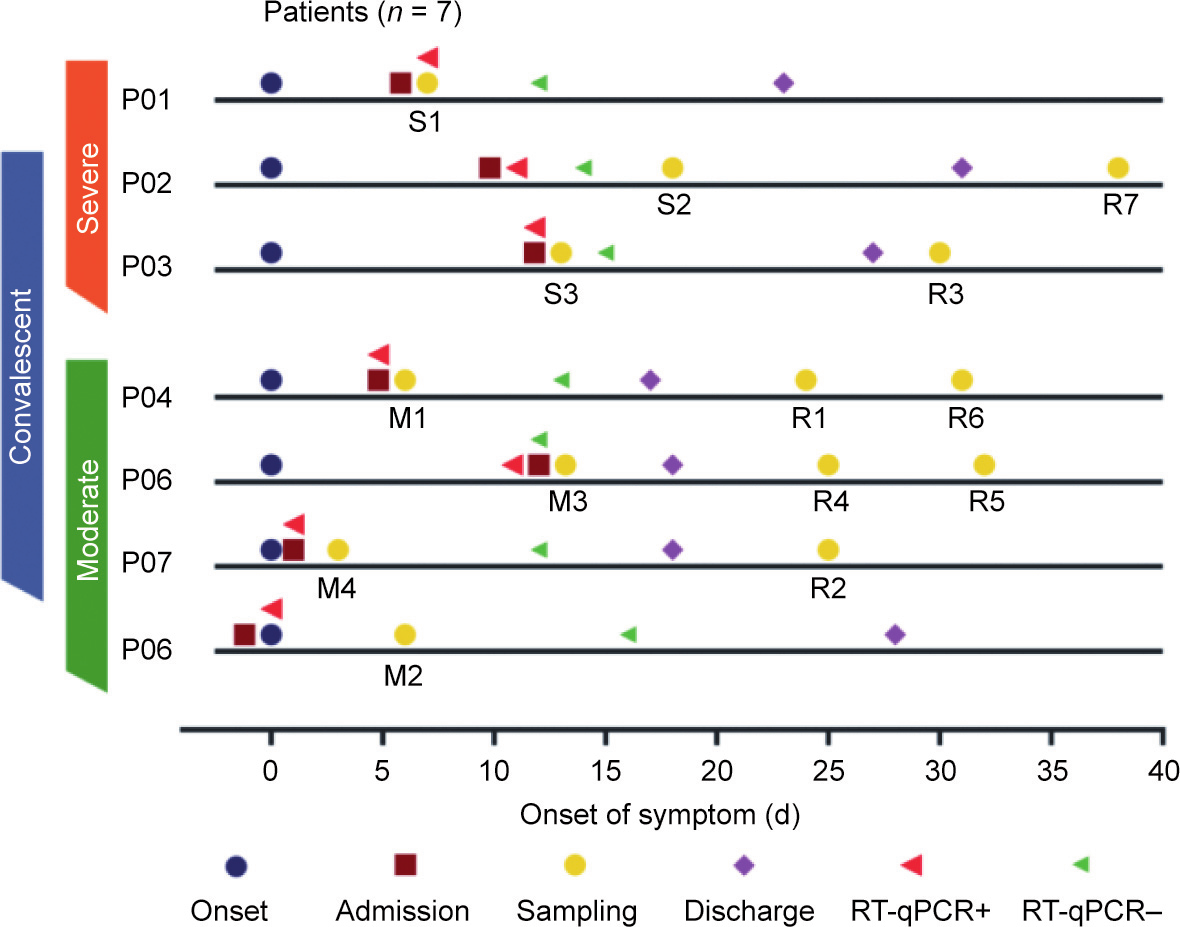
Fig. 1
Fig. 2
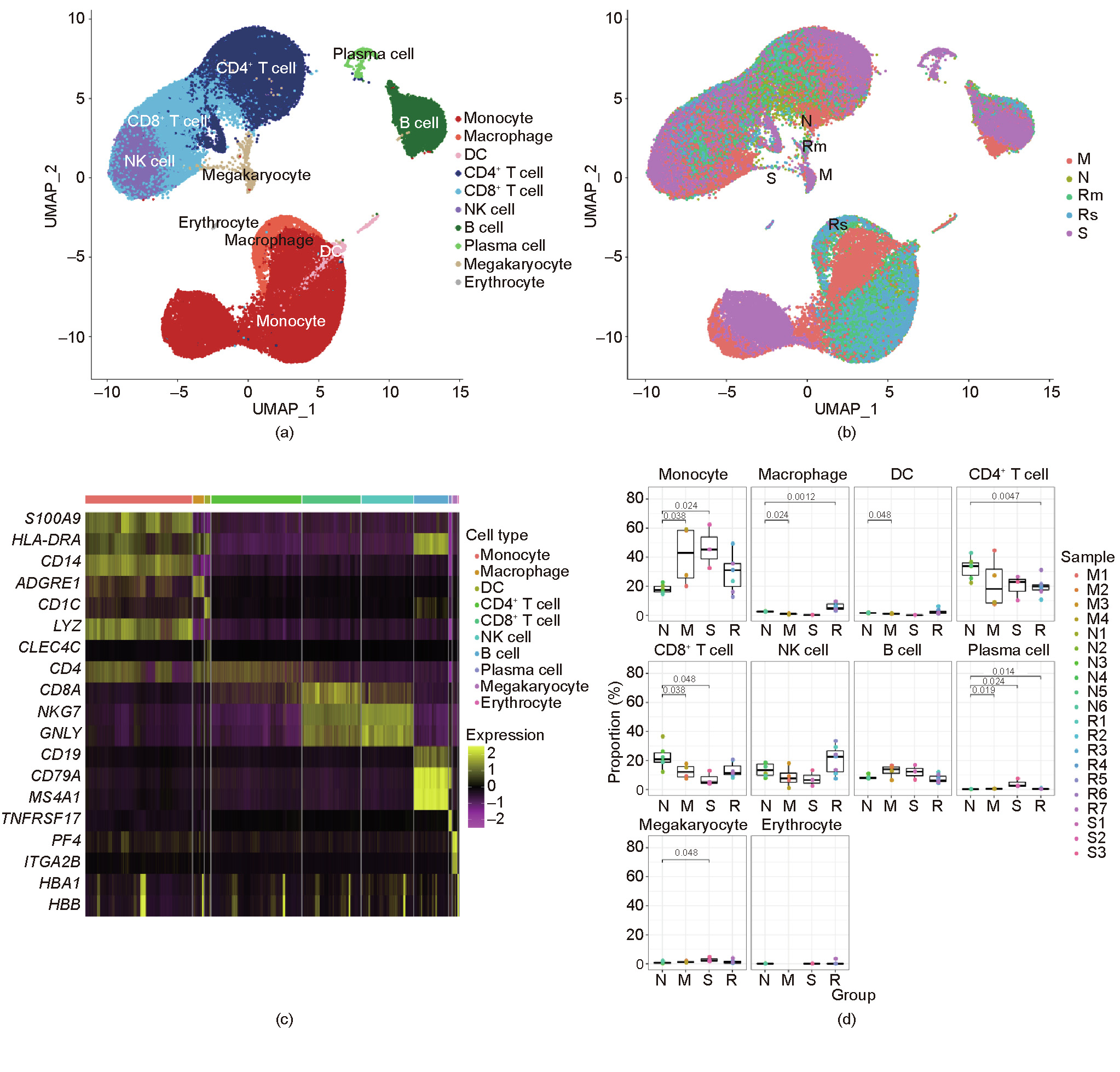
Fig. 3

Fig. 4

Fig. 5
References
[ 1 ] Zhang W, Zhao Y, Zhang F, Wang Q, Li T, Liu Z, et al. The use of antiinflammatory drugs in the treatment of people with severe coronavirus disease 2019 (COVID-19): the perspectives of clinical immunologists from China. Clin Immunol 2020;214:108393. link1
[ 2 ] Jafarzadeh A, Chauhan P, Saha B, Jafarzadeh S, Nemati M. Contribution of monocytes and macrophages to the local tissue inflammation and cytokine storm in COVID-19: lessons from SARS and MERS, and potential therapeutic interventions. Life Sci 2020;257:118102. link1
[ 3 ] Mehta P, McAuley DF, Brown M, Sanchez E, Tattersall RS, Manson JJ; HLH Across Speciality Collaboration, UK. COVID-19: consider cytokine storm syndromes and immunosuppression. Lancet 2020;395(10229):1033–4.
[ 4 ] Misharin AV, Morales-Nebreda L, Reyfman PA, Cuda CM, Walter JM, McQuattie-Pimentel AC, et al. Monocyte-derived alveolar macrophages drive lung fibrosis and persist in the lung over the life span. J Exp Med 2017;214 (8):2387–404. link1
[ 5 ] Kapellos TS, Bonaguro L, Gemünd I, Reusch N, Saglam A, Hinkley ER, et al. Human monocyte subsets and phenotypes in major chronic inflammatory diseases. Front Immunol 2019;10:2035. link1
[ 6 ] Yu M, Liu Y, Xu D, Zhang R, Lan L, Xu H. Prediction of the development of pulmonary fibrosis using serial thin-section CT and clinical features in patients discharged after treatment for COVID-19 pneumonia. Korean J Radiol 2020;21 (6):746–55. link1
[ 7 ] Han W, Zhu M, Chen J, Zhang J, Zhu S, Li T, et al. Lung transplantation for elderly patients with end-stage COVID-19 pneumonia. Ann Surg 2020;272(1): e33–4. link1
[ 8 ] Guillaumet-Adkins A, Rodríguez-Esteban G, Mereu E, Mendez-Lago M, Jaitin DA, Villanueva A, et al. Single-cell transcriptome conservation in cryopreserved cells and tissues. Genome Biol 2017;18(1):45. link1
[ 9 ] Aibar S, González-Blas CB, Moerman T, Huynh-Thu VA, Imrichova H, Hulselmans G, et al. SCENIC: single-cell regulatory network inference and clustering. Nat Methods 2017;14(11):1083–6. link1
[10] Weiss ES, Girard-Guyonvarc’h C, Holzinger D, de Jesus AA, Tariq Z, Picarsic J, et al. Interleukin-18 diagnostically distinguishes and pathogenically promotes human and murine macrophage activation syndrome. Blood 2018;131 (13):1442–55. link1
[11] Neufert C, Becker C, Türeci Ö, Waldner MJ, Backert I, Floh K, et al. Tumor fibroblast-derived epiregulin promotes growth of colitis-associated neoplasms through ERK. J Clin Invest 2013;123(4):1428–43. link1
[12] Zaiss DMW, Gause WC, Osborne LC, Artis D. Emerging functions of amphiregulin in orchestrating immunity, inflammation, and tissue repair. Immunity 2015;42(2):216–26. link1
[13] Venkataraman T, Frieman MB. The role of epidermal growth factor receptor (EGFR) signaling in SARS coronavirus-induced pulmonary fibrosis. Antiviral Res 2017;143:142–50. link1
[14] Chan KS, Zheng JP, Mok YW, Li YM, Liu YN, Chu CM, et al. SARS: prognosis, outcome and sequelae. Respirology 2003;8(S1):S36–40. link1
[15] Chan SY, Wong RW. Expression of epidermal growth factor in transgenic mice causes growth retardation. J Biol Chem 2000;275(49):38693–8. link1
[16] Ding L, Liu T, Wu Z, Hu B. Bone marrow CD11c+ cell-derived amphiregulin promotes pulmonary fibrosis. J Immunol 2016;197(1):303–12.
[17] Frieman M, Yount B, Agnihothram S, Page C, Donaldson E, Roberts A, et al. Molecular determinants of severe acute respiratory syndrome coronavirus pathogenesis and virulence in young and aged mouse models of human disease. J Virol 2012;86(2):884–97. link1
[18] Verweyen E, Holzinger D, Weinhage T, Hinze C. Synergistic signaling of TLR and IFNa/b facilitates escape of IL-18 expression from endotoxin tolerance. Am J Respir Crit Care Med 2020;201(5):526–39. link1
[19] Slaats J, Ten Oever J, van de Veerdonk FL, Netea MG. IL-1b/IL-6/CRP and IL18/ferritin: distinct inflammatory programs in infections. PLoS Pathog 2016;12 (12):e1005973. link1
[20] Hoshino T, Okamoto M, Sakazaki Y, Kato S, Young HA, Aizawa H. Role of proinflammatory cytokines IL-18 and IL-1b in bleomycin-induced lung injury in humans and mice. Am J Respir Cell Mol Biol 2009;41(6):661–70. link1
[21] Kang MJ, Choi JM, Kim BH, Lee CM, Cho WK, Choe G, et al. IL-18 induces emphysema and airway and vascular remodeling via IFN-c, IL-17A, and IL-13. Am J Respir Crit Care Med 2012;185(11):1205–17. link1
[22] Arend WP, Palmer G, Gabay C. IL-1, IL-18, and IL-33 families of cytokines. Immunol Rev 2008;223(1):20–38. link1
[23] Rodrigues TS, de Sá KSG, Ishimoto AY, Becerra A, Oliveira S, Almeida L, et al. Inflammasomes are activated in response to SARS-CoV-2 infection and are associated with COVID-19 severity in patients. J Exp Med 2021;218(3): e20201707. link1
[24] McGonagle D, Sharif K, O’Regan A, Bridgewood C. The role of cytokines including interleukin-6 in COVID-19 induced pneumonia and macrophage activation syndrome-like disease. Autoimmun Rev 2020;19(6):102537. link1
[25] Vecchié A, Bonaventura A. IL-18 and infections: is there a role for targeted therapies? J Cell Physiol 2021;236(3):1638–57. link1
[26] Lachmandas E, Boutens L, Ratter JM, Hijmans A, Hooiveld GJ. Microbial stimulation of different Toll-like receptor signalling pathways induces diverse metabolic programmes in human monocytes. Nat Microbiol 2016; 2:16246. link1
[27] Yamada KJ, Heim CE, Xi X. Monocyte metabolic reprogramming promotes proinflammatory activity and Staphylococcus aureus biofilm clearance. PLoS Pathog 2020;16(3):e1008354. link1
[28] Gibellini L, De Biasi S. Altered bioenergetics and mitochondrial dysfunction of monocytes in patients with COVID-19 pneumonia. EMBO Mol Med 2020;12 (12):e13001. link1
[29] Codo AC, Davanzo GG, Monteiro LB, de Souza GF, Muraro SP, Virgilio-da-Silva JV, et al. Elevated glucose levels favor SARS-CoV-2 infection and monocyte response through a HIF-1a/glycolysis-dependent axis. Cell Metab 2020;32 (3):437–46.e5.
[30] Ardestani A, Azizi Z. Targeting glucose metabolism for treatment of COVID-19. Signal Transduct Target Ther 2021;6(1):112. link1
[31] Barberis E, Timo S, Amede E, Vanella VV, Puricelli C. Large-scale plasma analysis revealed new mechanisms and molecules associated with the host response to SARS-CoV-2. Int J Mol Sci 2020;21(22):8623. link1
[32] Savage DB, Sewter CP, Klenk ES, Segal DG, Vidal-Puig A, Considine RV, et al. Resistin/Fizz3 expression in relation to obesity and peroxisome proliferator-activated receptor-c action in humans. Diabetes 2001;50 (10):2199–202. link1
[33] Tripathi D, Kant S, Pandey S. Resistin in metabolism, inflammation, and disease. FEBS J 2020;287(15):3141–9. link1
[34] Hemmat N, Derakhshani A, Bannazadeh Baghi H, Silvestris N, Baradaran B, de Summa S. Neutrophils, crucial, or harmful immune cells involved in coronavirus infection: a bioinformatics study. Front Genet 2020;11:641. link1
[35] Meizlish ML, Pine AB, Bishai JD, Goshua G, Nadelmann ER, Simonov M, et al. A neutrophil activation signature predicts critical illness and mortality in COVID-19. Blood Adv 2021;5(5):1164–77. link1
[36] McTernan CL, McTernan PG, Harte AL, Levick PL, Barnett AH, Kumar S. Resistin, central obesity, and type 2 diabetes. Lancet 2002;359(9300):46–7. link1
[37] Reilly MP, Lehrke M, Wolfe ML, Rohatgi A, Lazar MA, Rader DJ. Resistin is an inflammatory marker of atherosclerosis in humans. Circulation 2005;111 (7):932–9. link1











 京公网安备 11010502051620号
京公网安备 11010502051620号




