
2020, Volume 6, Issue 5
Engineering >> 2020, Volume 6, Issue 5 doi: 10.1016/j.eng.2019.07.027
Putative Mode of Action of the Monoterpenoids Linalool, Methyl Eugenol, Estragole, and Citronellal on Ligand-Gated Ion Channels
a College of Natural and Computational Sciences, Hawaii Pacific University, Kaneohe, HI 96744, USA
b Department of Molecular Biosciences and Bioengineering, University of Hawaii at Manoa, Honolulu, HI 96822, USA
c Department of Internal Medicine, University of Colorado School of Medicine, Aurora, CO 80045, USA
† Current address: Guangdong Provincial Key Laboratory of Sports and Health Promotion, Scientific Research Center, Guangzhou Sport University, Guangdong 510500, China
Next Previous
Abstract
Essential oil has been used as sedatives, anticonvulsants, and local anesthetics in traditional medical remedies; as preservatives for food, fruit, vegetable, and grain storage; and as bio-pesticides for food production. Linalool (LL), along with a few other major components such as methyl eugenol (ME), estragole (EG), and citronellal, are the active chemicals in many essential oils such as basil oil. Basil oil and the aforementioned monoterpenoids are potent against insect pests. However, the molecular mechanism of action of these chemical constituents is not well understood. It is well-known that the γ-aminobutyric acid type A receptors (GABAARs) and nicotinic acetylcholine receptor (nAChR) are primary molecular targets of the synthetic insecticides used in the market today. Furthermore, GABAAR-targeted therapeutics have been used in clinics for many decades, including barbiturates and benzodiazepines, to name just a few. In this research, we studied the electrophysiological effects of LL, ME, EG, and citronellal on GABAAR and nAChR to further understand their versatility as therapeutic agents in traditional remedies and as insecticides. Our results revealed that LL inhibits both GABAAR and nAChR, which may explain its insecticidal activity. LL is a concentration-dependent, non-competitive inhibitor on GABAAR, as the half-maximal effective concentration values of γ-aminobutyric acid (GABA) for the rat α1β3γ2L GABAAR were not affected by LL: (36.2 ± 7.9) µmol·L−1 and (36.1 ± 23.8) µmol·L−1 in the absence and presence of 5 mmol·L−1 LL, respectively. The half-maximal inhibitory concentration (IC50) of LL on GABAAR was approximately 3.2 mmol·L−1. Considering that multiple monoterpenoids are found within the same essential oil, it is likely that LL has a synergistic effect with ME, which has been previously characterized as both a GABAAR agonist and a positive allosteric modulator, and with other monoterpenoids, which offers a possible explanation for the sedative and anticonvulsant effects and the insecticidal activities of LL.
Keywords
Essential oil ; γ-aminobutyric acid type A receptor ; Linalool ; Monoterpenoid ; Nicotinic acetylcholine receptor
Figures

Fig. 1

Fig. 2
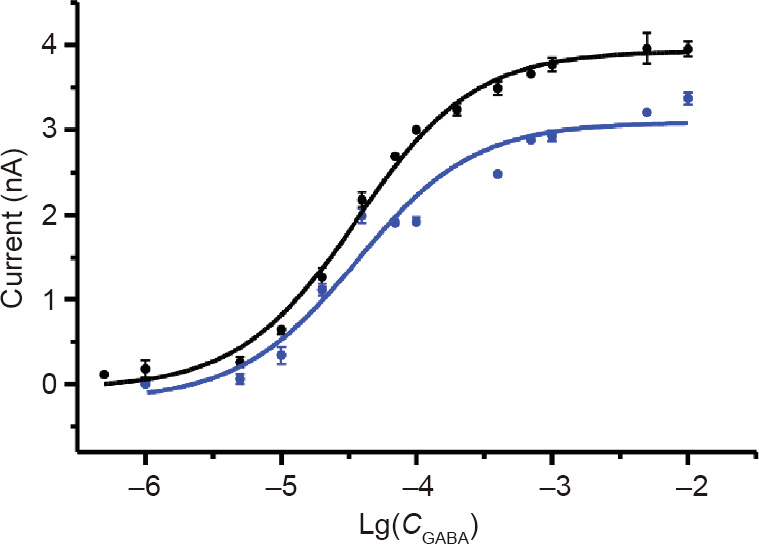
Fig. 3
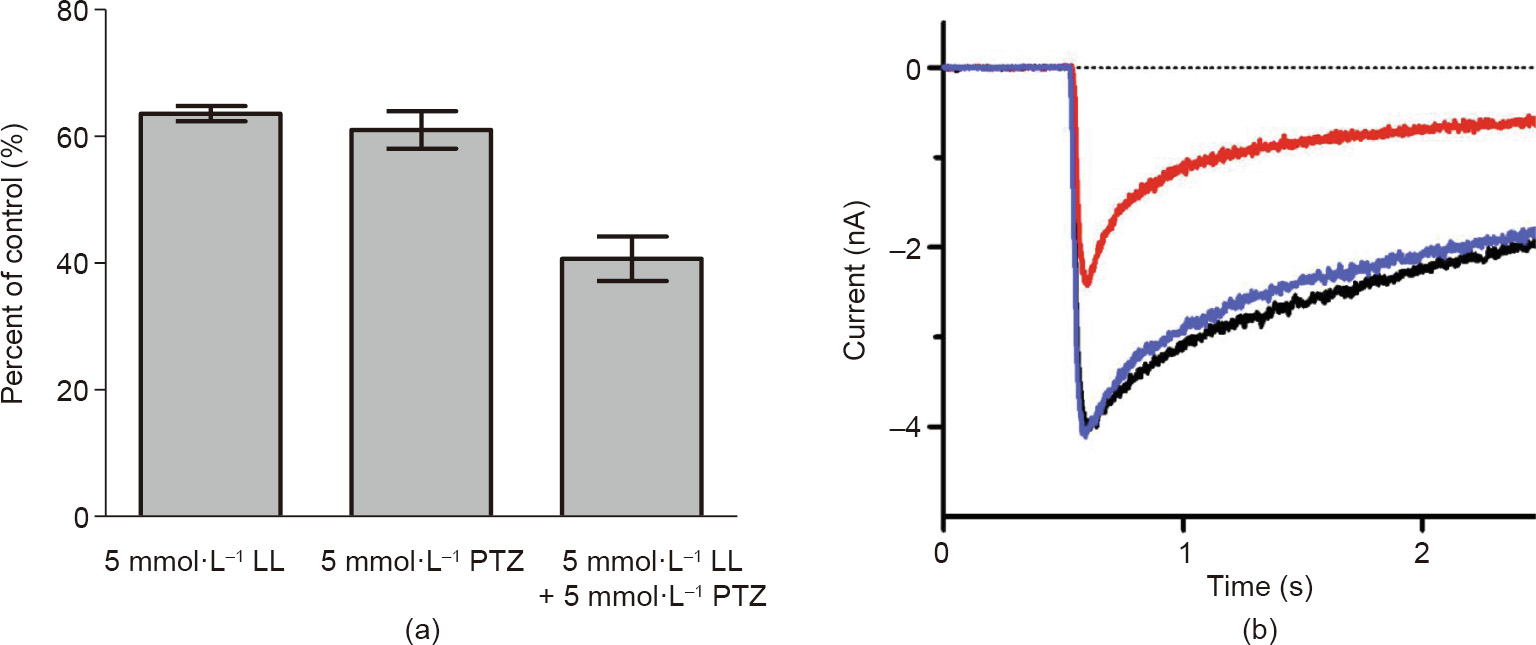
Fig. 4

Fig. 5
References
[ 1 ] Jeon S, Hur J, Jeong HJ, Koo BS, Pak SC. SuHeXiang Wan essential oil alleviates amyloid beta induced memory impairment through inhibition of tau protein phosphorylation in mice. Am J Chin Med 2011;39(5):917–32. link1
[ 2 ] Agra MF, Baracho GS, Nurit K, Basílio IJ, Coelho VP. Medicinal and poisonous diversity of the flora of ‘‘Cariri Paraibano”, Brazil. J Ethnopharmacol 2007;111 (2):383–95. link1
[ 3 ] Lawless J. The encyclopedia of essential oils: the complete guide to the use of aromatic oils in aromatherapy, herbalism, health, and well being. San Francisco: Conari Press; 2013. link1
[ 4 ] Preedy VR, editor. Essential oils in food preservation, flavor and safety. Amsterdam: Academic Press; 2016. link1
[ 5 ] Chang CL, Cho IK, Li QX. Insecticidal activity of basil oil, trans-anethole, estragole, and linalool to adult fruit flies of Ceratitis capitata, Bactrocera dorsalis, and Bactrocera cucurbitae. J Econ Entomol 2009;102(1):203–9. link1
[ 6 ] Jankowska M, Rogalska J, Wyszkowska J, Stankiewicz M. Molecular targets for components of essential oils in the insect nervous system—a review. Molecules 2017;23(1):34. link1
[ 7 ] Letizia CS, Cocchiara J, Lalko J, Api AM. Fragrance material review on linalool. Food Chem Toxicol 2003;41(7):943–64. link1
[ 8 ] de Almeida RN, Agra Mde F, Maior FN, de Sousa DP. Essential oils and their constituents: anticonvulsant activity. Molecules 2011;16(3):2726–42. link1
[ 9 ] Nascimento SS, Araújo AA, Brito RG, Serafini MR, Menezes PP, DeSantana JM, et al. Cyclodextrin-complexed Ocimum basilicum leaves essential oil increases Fos protein expression in the central nervous system and produce an antihyperalgesic effect in animal models for fibromyalgia. Int J Mol Sci 2015;16(1):547–63. link1
[10] De Sousa DP, Nóbrega FF, Santos CC, de Almeida RN. Anticonvulsant activity of the linalool enantiomers and racemate: investigation of chiral influence. Nat Prod Commun 2010;5(12):1847–51. link1
[11] Batista PA, Werner MF, Oliveira EC, Burgos L, Pereira P, Brum LF, et al. Evidence for the involvement of ionotropic glutamatergic receptors on the antinociceptive effect of ()-linalool in mice. Neurosci Lett 2008;440 (3):299–303. link1
[12] Hirai M, Ito M. Sedative effects of the essential oil and headspace air of Ocimum basilicum by inhalation in mice. J Nat Med 2019;73(1):283–8. link1
[13] Melo FH, Moura BA, de Sousa DP, de Vasconcelos SM, Macedo DS, Fonteles MM, et al. Antidepressant-like effect of carvacrol (5-isopropyl-2- methylphenol) in mice: involvement of dopaminergic system. Fundam Clin Pharmacol 2011;25(3):362–7. link1
[14] Melo FH, Venâncio ET, de Sousa DP, de França Fonteles MM, de Vasconcelos SM, Viana GS, et al. Anxiolytic-like effect of carvacrol (5-isopropyl-2- methylphenol) in mice: involvement with GABAergic transmission. Fundam Clin Pharmacol 2010;24(4):437–43. link1
[15] Peana AT, De Montis MG, Sechi S, Sircana G, D’Aquila PS, Pippia P. Effects of ()-linalool in the acute hyperalgesia induced by carrageenan, L-glutamate and prostaglandin E2. Eur J Pharmacol 2004;497(3):279–84. link1
[16] Malcolm BJ, Tallian K. Essential oil of lavender in anxiety disorders: ready for prime time? Ment Health Clin 2018;7(4):147–55. link1
[17] Greenfield LJ Jr. Molecular mechanisms of antiseizure drug activity at GABAA receptors. Seizure 2013;22(8):589–600. link1
[18] Kamin´ ski K, Obniska J, Chlebek I, Liana P, Pe˛kala E. Synthesis and biological properties of new N-Mannich bases derived from 3-methyl-3-phenyl- and 3,3- dimethyl-succinimides. Part V Eur J Med Chem 2013;66:12–21. link1
[19] Zheng C, Yang K, Liu Q, Wang MY, Shen J, Vallés AS, et al. The anticonvulsive drug lamotrigine blocks neuronal a4b2 nicotinic acetylcholine receptors. J Pharmacol Exp Ther 2010;335(2):401–8. link1
[20] Xie X, Lancaster B, Peakman T, Garthwaite J. Interaction of the antiepileptic drug lamotrigine with recombinant rat brain type IIA Na+ channels and with native Na+ channels in rat hippocampal neurones. Pflugers Arch 1995;430 (3):437–46. link1
[21] Li QX, Chang CL. Basil (Ocimum basilicum L.) oils. In: Preedy VR, editor. Essential oils in food preservation, flavor and safety. Amsterdam: Academic Press; 2016. p. 231–8. link1
[22] Wong G, Sei Y, Skolnick P. Stable expression of type I gamma-aminobutyric acid-A/benzodiazepine receptors in a transfected cell line. Mol Pharmacol 1992;42(6):996–1003. link1
[23] Davies PA, Hoffmann EB, Carlisle HJ, Tyndale RF, Hales TG. The influence of an endogenous b3 subunit on recombinant GABAA receptor assembly and pharmacology in WSS-1 cells and transiently transfected HEK293 cells. Neuropharmacology 2000;39(4):611–20. link1
[24] Xiao Y, Meyer EL, Thompson JM, Surin A, Wroblewski J, Kellar KJ. Rat a3/b4 subtype of neuronal nicotinic acetylcholine receptor stably expressed in a transfected cell line: pharmacology of ligand binding and function. Mol Pharmacol 1998;54(2):322–33. link1
[25] Udgaonkar JB, Hess GP. Chemical kinetic measurements of a mammalian acetylcholine receptor by a fast-reaction technique. Proc Natl Acad Sci USA 1987;84(24):8758–62. link1
[26] Akk G, Steinbach JH. Activation and block of recombinant GABAA receptors by pentobarbitone: a single-channel study. Br J Pharmacol 2000;130 (2):249–58. link1
[27] Steinbach JH, Akk G. Modulation of GABAA receptor channel gating by pentobarbital. J Physiol 2001;537(Pt 3):715–33. link1
[28] Dibas MI, Dillon GH. The central nervous system convulsant pentylenetetrazole stimulates gamma-aminobutyric acid (GABA)-activated current in picrotoxin-resistant GABAA receptors in HEK293 cells. Neurosci Lett 2000;285(3):193–6. link1
[29] Gurley D, Amin J, Ross PC, Weiss DS, White G. Point mutations in the M2 region of the alpha, beta, or gamma subunit of the GABAA channel that abolish block by picrotoxin. Receptor Channel 1995;3(1):13–20. link1
[30] Ding J, Huang C, Peng Z, Xie Y, Deng S, Nie YZ, et al. Electrophysiological characterization of methyleugenol: a novel agonist of GABAA receptors. ACS Chem Neurosci 2014;5(9):803–11. link1
[31] Melo MS, Sena LC, Barreto FJN, Bonjardim LR, Almeida JRGS, Lima JT, et al. Antinociceptive effect of citronellal in mice. Pharm Biol 2010;48(4):411–6. link1
[32] de Sousa DP, Gonçalves JCR, Quintans-Júnior L, Cruz JS, Araújo DA, de Almeida RN. Study of anticonvulsant effect of citronellol, a monoterpene alcohol, in rodents. Neurosci Lett 2006;401(3):231–5. link1
[33] Elisabetsky E, Brum LFS, Souza DO. Anticonvulsant properties of linalool in glutamate-related seizure models. Phytomedicine 1999;6(2):107–13. link1
[34] Narusuye K, Kawai F, Matsuzaki K, Miyachi E. Linalool suppresses voltagegated currents in sensory neurons and cerebellar Purkinje cells. J Neural Transm (Vienna) 2005;112(2):193–203. link1
[35] Leal-Cardoso JH, da Silva-Alves KS, Ferreira-da-Silva FW, dos SantosNascimento T, Joca HC, de Macedo FH, et al. Linalool blocks excitability in peripheral nerves and voltage-dependent Na+ current in dissociated dorsal root ganglia neurons. Eur J Pharmacol 2010;645(1–3):86–93. link1
[36] Brum LF, Elisabetsky E, Souza D. Effects of linalool on [3 H] MK801 and [3 H] muscimol binding in mouse cortical membranes. Phytother Res 2001;15 (5):422–5. link1
[37] Re L, Barocci S, Sonnino S, Mencarelli A, Vivani C, Paolucci G, et al. Linalool modifies the nicotinic receptor-ion channel kinetics at the mouse neuromuscular junction. Pharmacol Res 2000;42(2):177–82. link1
[38] Albuquerque EX, Pereira EF, Alkondon M, Rogers SW. Mammalian nicotinic acetylcholine receptors: from structure to function. Physiol Rev 2009;89 (1):73–120. link1
[39] Rogawski MA, Löscher W. The neurobiology of antiepileptic drugs. Nat Rev Neurosci 2004;5(7):553–64. link1
[40] Cong D, Tang Z, Li L, Huang Y, Wang J, Chen L. Cross-talk between NMDA and GABAA receptors in cultured neurons of the rat inferior colliculus. Sci China Life Sci 2011;54(6):560–6. link1
[41] Chisari M, Zorumski CF, Mennerick S. Cross talk between synaptic receptors mediates NMDA-induced suppression of inhibition. J Neurophysiol 2012;107 (9):2532–40. link1
[42] Chen QX, Stelzer A, Kay AR, Wong RK. GABAA receptor function is regulated by phosphorylation in acutely dissociated guinea-pig hippocampal neurones. J Physiol 1990;420(1):207–21. link1
[43] Chen QX, Wong RK. Suppression of GABAA receptor responses by NMDA application in hippocampal neurones acutely isolated from the adult guineapig. J Physiol 1995;482(Pt 2):353–62. link1
[44] Zhu WJ, Vicini S, Harris BT, Grayson DR. NMDA-mediated modulation of gamma-aminobutyric acid type A receptor function in cerebellar granule neurons. J Neurosci 1995;15(11):7692–701. link1
[45] Stelzer A, Slater NT, ten Bruggencate G. Activation of NMDA receptors blocks GABAergic inhibition in an in vitro model of epilepsy. Nature 1987;326 (6114):698–701. link1
[46] Zhang J, Berg DK. Reversible inhibition of GABAA receptors by a7-containing nicotinic receptors on the vertebrate postsynaptic neurons. J Physiol 2007;579 (Pt 3):753–63. link1











 京公网安备 11010502051620号
京公网安备 11010502051620号




