
2020, Volume 6, Issue 1
Engineering >> 2020, Volume 6, Issue 1 doi: 10.1016/j.eng.2019.08.014
First Isolation and Characterization of Ochrobactrum anthropi from Pig
a National Research Center for Veterinary Medicine, Luoyang 471000, China
b College of Animal Science and Veterinary Medicine, Henan Agricultural University, Zhengzhou 450002, China
# These authors contributed equally to this work.
Next Previous
Abstract
One Gram-negative bacillus was isolated from a brain sample of a pig with neurological symptoms. Pathological examination showed meningitis at necropsy. Ochrobactrum anthropi (O. anthropi) was successfully isolated from the brain sample and was confirmed by biochemical reaction results (API 20 NE) and gene sequencing. The strain was highly resistant to beta-lactam antibiotics. Mice were experimentally infected with O. anthropi and showed typical meningitis. This is the first report on O. anthropi isolated from a pig, and indicates that O. anthropi may have a broader host spectrum of infection.
Keywords
Figures

Fig. 1

Fig. 2

Fig. 3
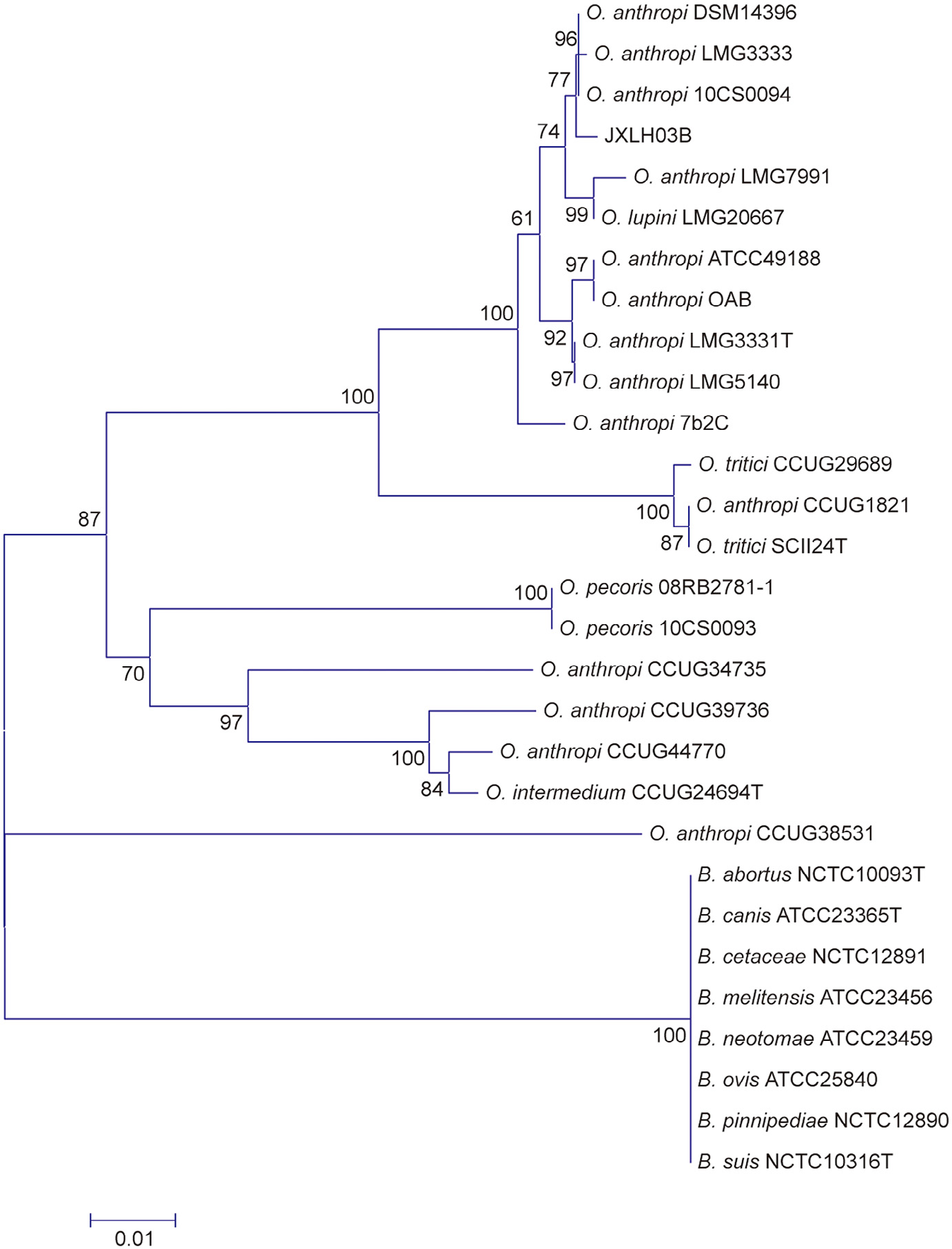
Fig. 4
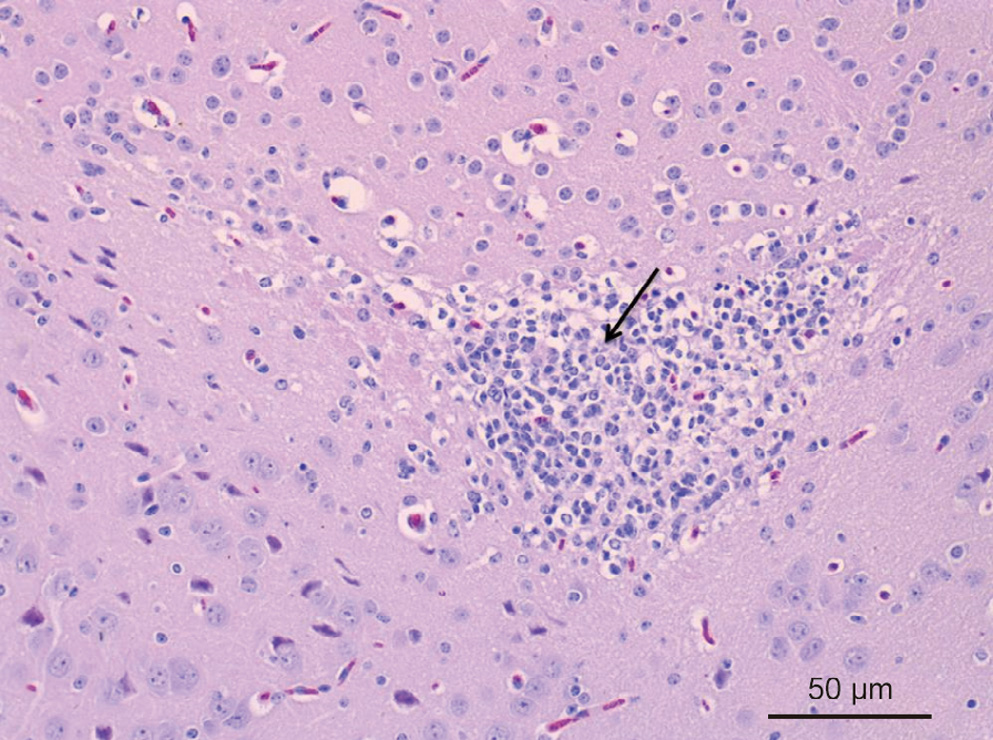
Fig. 5
References
[ 1 ] Holmes B, Popoff M, Kiredjian M, Kersters K. Ochrobactrum anthropi gen. nov., sp. nov. from human clinical specimens and previously known as group Vd. Int J Syst Bacteriol 1988;38(4):406–16. link1
[ 2 ] Lebuhn M, Achouak W, Schloter M, Berge O, Meier H, Barakat M, et al. Taxonomic characterization of Ochrobactrum sp. isolates from soil samples and wheat roots, and description of Ochrobactrum tritici sp. nov. and Ochrobactrum grignonense sp. nov. Int J Syst Evol Microbiol 2000;50(Pt 6):2207–23. link1
[ 3 ] Alnor D, Frimodt-Møller N, Espersen F, Frederiksen W. Infections with the unusual human pathogens Agrobacterium species and Ochrobactrum anthropi. Clin Infect Dis 1994;18(6):914–20. link1
[ 4 ] Cieslak TJ, Robb ML, Drabick CJ, Fischer GW. Catheter-associated sepsis caused by Ochrobactrum anthropi: report of a case and review of related nonfermentative bacteria. Clin Infect Dis 1992;14(4):902–7. link1
[ 5 ] Berman AJ, Del Priore LV, Fischer CK. Endogenous Ochrobactrum anthropi endophthalmitis. Am J Ophthalmol 1997;123(4):560–2. link1
[ 6 ] Braun M, Jonas JB, Schönherr U, Naumann GO. Ochrobactrum anthropi endophthalmitis after uncomplicated cataract surgery. Am J Ophthalmol 1996;122(2):272–3. link1
[ 7 ] Galanakis E, Bitsori M, Samonis G, Christidou A, Georgiladakis A, Sbyrakis S, et al. Ochrobactrum anthropi bacteraemia in immunocompetent children. Scand J Infect Dis 2002;34(11):800–3. link1
[ 8 ] Romero Gómez MP, Peinado Esteban AM, Sobrino Daza JA, Sáez Nieto JA, Alvarez D, Peña García P. Prosthetic mitral valve endocarditis due to Ochrobactrum anthropi: case report. J Clin Microbiol 2004;42(7):3371–3. link1
[ 9 ] Appelbaum PC, Campbell DB. Pancreatic abscess associated with achromobacter group Vd biovar 1. J Clin Microbiol 1980;12(2):282–3. link1
[10] Barson WJ, Cromer BA, Marcon MJ. Puncture wound osteochondritis of the foot caused by CDC group Vd. J Clin Microbiol 1987;25(10):2014–6. link1
[11] Chang HJ, Christenson JC, Pavia AT, Bobrin BD, Bland LA, Carson LA, et al. Ochrobactrum anthropi meningitis in pediatric pericardial allograft transplant recipients. J Infect Dis 1996;173(3):656–60. link1
[12] Christenson JC, Pavia AT, Seskin K, Brockmeyer D, Korgenski EK, Jenkins E, et al. Meningitis due to Ochrobactrum anthropi: an emerging nosocomial pathogen. A report of three cases. Pediatr Neurosurg 1997;27(4):218–21. link1
[13] Vaidya SA, Citron DM, Fine MB, Murakami G, Goldstein EJ. Pelvic abscess due to Ochrobactrum intermedium [corrected] in an immunocompetent host: case report and review of the literature. J Clin Microbiol 2006;44(3):1184–6. link1
[14] Jelveh N, Cunha BA. Ochrobactrum anthropi bacteremia. Heart Lung 1999;28 (2):145–6. link1
[15] Franci P, Dotto G, Cattai A, Pasotto D. Lethal septic shock after dental scaling in a healthy dog due to Ochrobactrum anthropi-contaminated propofol. J Small Anim Pract 2015;56(5):345–7. link1
[16] El-Adawy H, Hotzel H, Tomaso H, Neubauer H, Hafez HM. Isolation and characterization of Ochrobactrum anthropi and Ochrobactrum pecoris from caecal content of commercial turkeys. Vet Microbiol 2012;155(2–4):349–54. link1
[17] Gerhardt P, Murray RGE, Wood WA, Krieg NR, editors. Methods for general and molecular bacteriology. Washington, DC: American Society for Microbiology; 1994. link1
[18] Scholz HC, Tomaso H, Al Dahouk S, Witte A, Schloter M, Kämpfer P, et al. Genotyping of Ochrobactrum anthropi by recA-based comparative sequence, PCR-RFLP, and 16S rRNA gene analysis. FEMS Microbiol Lett 2006;257(1):7–16. link1
[19] Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG. The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res 1997;25 (24):4876–82. link1
[20] Kimura M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J Mol Evol 1980;16(2):111–20. link1
[21] Saitou N, Nei M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol 1987;4(4):406–25. link1
[22] Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 2011;28(10):2731–9. link1
[23] Felsenstein J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution 1985;39(4):783–91. link1
[24] Bauer AW, Kirby WM, Sherris JC, Turck M. Antibiotic susceptibility testing by a standardized single disk method. Am J Clin Pathol 1966;45(4):493–6. link1
[25] Clinical and Laboratory Standards Institute. Performance standards for antimicrobial disk and dilution susceptibility tests for bacteria isolated from animals: approved standard. 4th ed. Wayne: National Committee for Clinical Laboratory Standards; 2013. link1
[26] Velasco J, Romero C, López-Goñi I, Leiva J, Díaz R, Moriyón I. Evaluation of the relatedness of Brucella spp. and Ochrobactrum anthropi and description of Ochrobactrum intermedium sp. nov., a new species with a closer relationship to Brucella spp. Int J Syst Bacteriol 1998;48(Pt 3):759–68. link1
[27] Kämpfer P, Citron DM, Goldstein EJ, Scholz HC. Difficulty in the identification and differentiation of clinically relevant Ochrobactrum species. J Med Microbiol 2007;56(Pt 11):1571–3. link1
[28] Renukaradhya GJ, Alekseev K, Jung K, Fang Y, Saif LJ. Porcine reproductive and respiratory syndrome virus-induced immunosuppression exacerbates the inflammatory response to porcine respiratory coronavirus in pigs. Viral Immunol 2010;23(5):457–66. link1
[29] Higgins CS, Avison MB, Jamieson L, Simm AM, Bennett PM, Walsh TR. Characterization, cloning and sequence analysis of the inducible Ochrobactrum anthropi AmpC beta-lactamase. J Antimicrob Chemother 2001;47(6):745–54. link1
[30] Oliver JW, Stapenhorst D, Warraich I, Griswold JA. Ochrobactrum anthropi and Delftia acidovorans to bacteremia in a patient with a gunshot wound. Infect Dis Clin Pract 2005;13(2):78–81. link1











 京公网安备 11010502051620号
京公网安备 11010502051620号




