
2020, Volume 6, Issue 1
Engineering >> 2020, Volume 6, Issue 1 doi: 10.1016/j.eng.2019.10.013
Presence and Antimicrobial Susceptibility of RE-cmeABC-Positive Campylobacter Isolated from Food-Producing Animals, 2014–2016
Beijing Advanced Innovation Center for Food Nutrition and Human Health, College of Veterinary Medicine, China Agricultural
University, Beijing 100193, China
Next Previous
Abstract
Campylobacter spp. (C. spp.) are the leading cause of human gastroenteritis worldwide. RE-CmeABC is a newly identified resistance-enhancing multidrug efflux pump of C. spp. that confers high-level resistance to fluoroquinolones, phenicols, macrolides, and tetracyclines, all of which are critical drugs in both human and veterinary medicine. In this study, we analyzed the presence and antimicrobial susceptibility of RE-cmeABC-positive Campylobacter isolates of food-animal origin from three representative regions (Shandong, Shanghai, and Guangdong) in China over three successive years, from 2014 to 2016. A total of 1088 Campylobacter isolates (931 C. coli and 157 C. jejuni) were recovered from the RE-cmeABC screening. We detected 122 (11.2%) RE-cmeABC-positive isolates of chicken origin, including 111 (70.7%) C. jejuni and 11 (1.2%) C. coli. This multidrug efflux pump is more prevalent among C. jejuni than C. coli. The level of resistance was significantly different in 111 RE-cmeABC-positive C. jejuni versus 46 RE-cmeABC-negative C. jejuni for florfenicol, clindamycin, and erythromycin (P < 0.05), but not for ciprofloxacin (CIP), tetracycline (TET), and gentamicin. However, the isolates harboring RE-cmeABC could shift the minimum inhibitory concentration distribution to the higher range for CIP and TET. Pulsed-field gel electrophoresis (PFGE) analysis suggested that horizontal transmission might be involved in the dissemination of RE-cmeABC in Shanghai and Guangdong, while clonal expansion was predominant in Shandong. Three isolates shared the indiscriminate PFGE types of RE-cmeABC-positive C. jejuni isolates in Shanghai and Guangdong, and four isolates in Shanghai and Shandong. Our study suggests the possibility of a wide dissemination of RE-cmeABC in Campylobacter of food-animal origin, which would pose a significant threat to public health.
Keywords
Figures
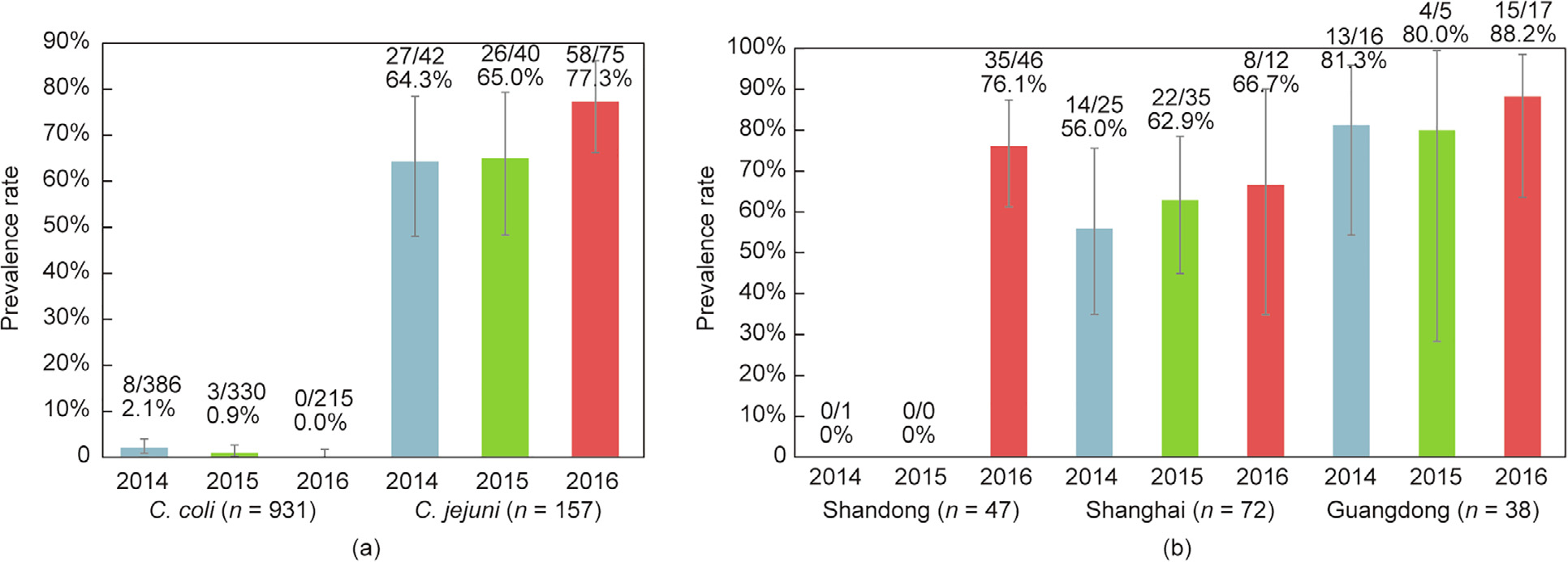
Fig. 1
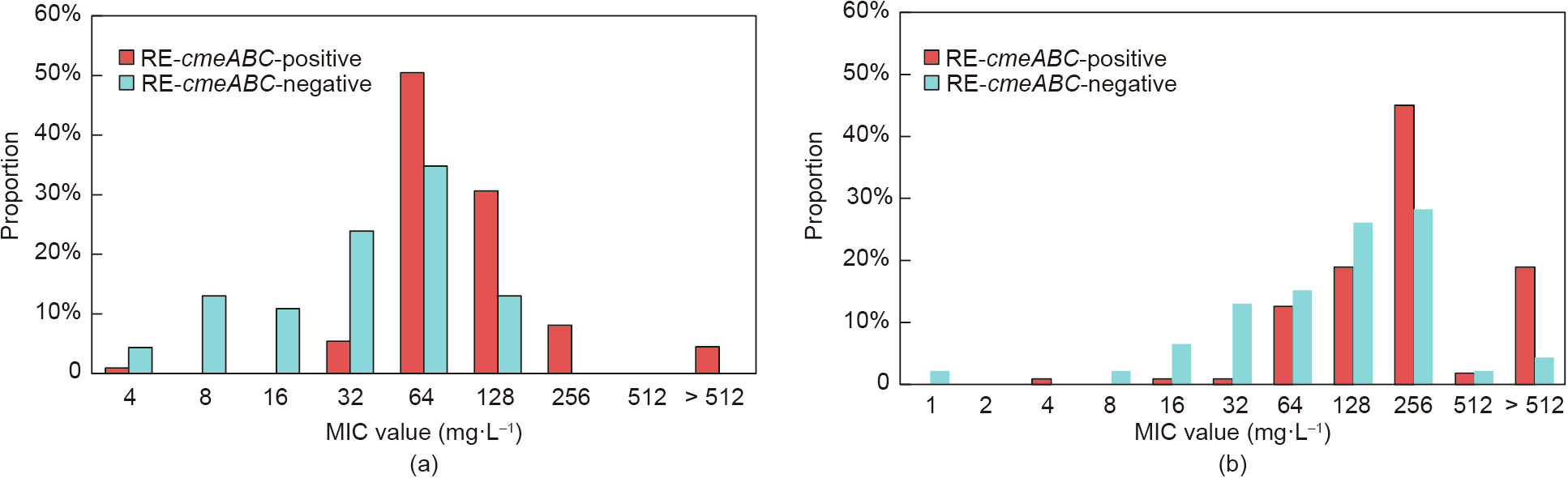
Fig. 2

Fig. 3
References
[ 1 ] Scallan E, Hoekstra RM, Angulo FJ, Tauxe RV, Widdowson MA, Roy SL, et al. Foodborne illness acquired in the United States—major pathogens. Emerg Infect Dis 2011;17:7–15. link1
[ 2 ] Kaakoush NO, Castano-Rodriguez N, Mitchell HM, Man SM. Global epidemiology of Campylobacter infection. Clin Microbiol Rev 2015;28:687–720. link1
[ 3 ] Man SM. The clinical importance of emerging Campylobacter species. Nat Rev Gastroenterol Hepatol 2011;8:669–85. link1
[ 4 ] Taylor EV, Herman KM, Ailes EC, Fitzgerald C, Yoder JS, Mahon BE, et al. Common source outbreaks of Campylobacter infection in the USA, 1997–2008. Epidemiol Infect 2013;141:987–96. link1
[ 5 ] Adak GK, Cowden JM, Nicholas S, Evans HS. The Public Health Laboratory Service national case-control study of primary indigenous sporadic cases of Campylobacter infection. Epidemiol Infect 1995;115:15–22. link1
[ 6 ] Allos BM. Campylobacter jejuni infections: update on emerging issues and trends. Clin Infect Dis 2001;32:1201–6. link1
[ 7 ] Luangtongkum T, Jeon B, Han J, Plummer P, Logue CM, Zhang Q. Antibiotic resistance in Campylobacter: emergence, transmission and persistence. Future Microbiol 2009;4:189–200. link1
[ 8 ] European Food Safety Authority, European Centre for Disease Prevention and Control. The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2017. EFSA J 2019;17(2):e05598. link1
[ 9 ] US Food and Drug Administration. The National Antimicrobial Resistance Monitoring System: NARMS integrated report, 2015. Report. Washington, DC: US Department of Health and Human Services, Food and Drug Administration; 2015.
[10] US Centers for Disease Control and Prevention. Antibiotic resistance threats in the United States, 2013 (AR threats report). Report. Washington, DC: US Department of Health and Human Services, Centers for Disease Control and Prevention; 2013.
[11] World Health Organization. Prioritization of pathogens to guide research and development of new antibiotics. Report. Geneva: World Health Organization; 2017.
[12] Zeng D, Zhang X, Xue F, Wang Y, Jiang L, Jiang Y. Phenotypic characters and molecular epidemiology of Campylobacter jejuni in East China. J Food Sci 2016;81:M106–13. link1
[13] Li Y, Zhang S, He M, Zhang Y, Fu Y, Liang H, et al. Prevalence and molecular characterization of Campylobacter spp. isolated from patients with diarrhea in Shunyi, Beijing. Front Microbiol 2018;9:52. link1
[14] Ju CY, Zhang MJ, Ma YP, Lu JR, Yu MH, Chen H, et al. Genetic and antibiotic resistance characteristics of Campylobacter jejuni isolated from diarrheal patients, poultry and cattle in Shenzhen. Biomed Environ Sci 2018;31:579–85. link1
[15] Du Y, Wang C, Ye Y, Liu Y, Wang A, Li Y, et al. Molecular identification of multidrug-resistant Campylobacter species from diarrheal patients and poultry meat in Shanghai, China. Front Microbiol 2018;9:1642. link1
[16] Wang Y, Dong Y, Deng F, Liu D, Yao H, Zhang Q, et al. Species shift and multidrug resistance of Campylobacter from chicken and swine, China, 2008– 14. J Antimicrob Chemother 2016;71:666–9. link1
[17] Payot S, Bolla JM, Corcoran D, Fanning S, Megraud F, Zhang Q. Mechanisms of fluoroquinolone and macrolide resistance in Campylobacter spp. Microbes Infect 2006;8:1967–71. link1
[18] Luo N, Sahin O, Lin J, Michel LO, Zhang Q. In vivo selection of Campylobacter isolates with high levels of fluoroquinolone resistance associated with gyrA mutations and the function of the CmeABC efflux pump. Antimicrob Agents Chemother 2003;47:390–4. link1
[19] Lin J, Michel LO, Zhang Q. CmeABC functions as a multidrug efflux system in Campylobacter jejuni. Antimicrob Agents Chemother 2002;46:2124–31. link1
[20] Pumbwe L, Piddock LJ. Identification and molecular characterisation of CmeB, a Campylobacter jejuni multidrug efflux pump. FEMS Microbiol Lett 2002;206:185–9. link1
[21] Yao H, Shen Z, Wang Y, Deng F, Liu D, Naren G, et al. Emergence of a potent multidrug efflux pump variant that enhances Campylobacter resistance to multiple antibiotics. mBio 2016;7(5):e01543–16. link1
[22] Cui M, Wu C, Zhang P, Wu C. Development of multiplex-mismatch amplification mutation-PCR assay for simultaneous detection of Campylobacter jejuni and mutation in gyrA gene related to fluoroquinolone resistance. Foodborne Pathog Dis 2016;13:642–5. link1
[23] Clinical and Laboratory Standards Institute. Methods for antimicrobial dilution and disk susceptibility testing of infrequently isolated and fastidious bacteria, 3rd edition. Document. Wayne: Clinical and Laboratory Standards Institute; 2016. Document No.: CLSI document M45.
[24] US Centers for Disease Control and Prevention. Antibiotics tested by NARMS [Internet]. Washington, DC: US Centers for Disease Control and Prevention [updated 2019 Mar 15; cited 2019 Aug 30]. Available from: https://www. cdc.gov/narms/antibiotics-tested.html/. link1
[25] Ribot EM, Fitzgerald C, Kubota K, Swaminathan B, Barrett TJ. Rapid pulsed-field gel electrophoresis protocol for subtyping of Campylobacter jejuni. J Clin Microbiol 2001;39:1889–94. link1
[26] Luangtongkum T, Morishita TY, Ison AJ, Huang S, McDermott PF. Zhang Q. Effect of conventional and organic production practices on the prevalence and antimicrobial resistance of Campylobacter spp. in poultry. Appl Environ Microbiol 2006;72:3600–7. link1
[27] Schwarz S, Kehrenberg C, Doublet B, Cloeckaert A. Molecular basis of bacterial resistance to chloramphenicol and florfenicol. FEMS Microbiol Rev 2004;28:519–42. link1
[28] Cagliero C, Mouline C, Payot S, Cloeckaert A. Involvement of the CmeABC efflux pump in the macrolide resistance of Campylobacter coli. J Antimicrob Chemother 2005;56:948–50. link1
[29] Lin J, Akiba M, Sahin O, Zhang QJ. CmeR functions as a transcriptional repressor for the multidrug efflux pump CmeABC in Campylobacter jejuni. Antimicrob Agents Chemother 2005;49:1067–75. link1











 京公网安备 11010502051620号
京公网安备 11010502051620号




