

《1. 引言》
1. 引言
临床中,一般建议有肾脏和肝脏疾病的患者减少蛋白质的摄入量。此外,由于饲料资源有限,为了节约饲料成本,低蛋白日粮在畜牧业中也得到了广泛的应用。但是研究表明,低蛋白饮食或日粮会降低人和动物大部分组织中的蛋白质沉积,因此可能影响患者健康[1]和动物的生长[2]。当蛋白质缺乏时,由于用于合成肌肉蛋白质的氨基酸的含量减少,造成肌肉萎缩[2–4]。
亮氨酸是10种必需氨基酸之一,可以调节人和动物蛋白质代谢,尤其是在餐后可以促进肌肉蛋白质合成,抑制蛋白质降解[5]。这可能是因为亮氨酸能够激活mTOR信号通路促进细胞内的翻译过程,包括促进S6K1和4E-BP1的磷酸化[6–8]。研究表明,长期低蛋白日粮处理会通过抑制mTOR信号通路减少蛋白质合成[2,9]。本文假设在低蛋白日粮中添加亮氨酸可以促进大鼠肌肉蛋白质合成,并保持骨骼肌重量。通过在低蛋白日粮中添加亮氨酸,探究其对大鼠骨骼肌重量及蛋白质合成的影响,并对相关机理(4E-BP1和S6K1的表达[10])进行探讨。
《2. 材料与方法》
2. 材料与方法
《2.1. 试验动物与日粮》
2.1. 试验动物与日粮
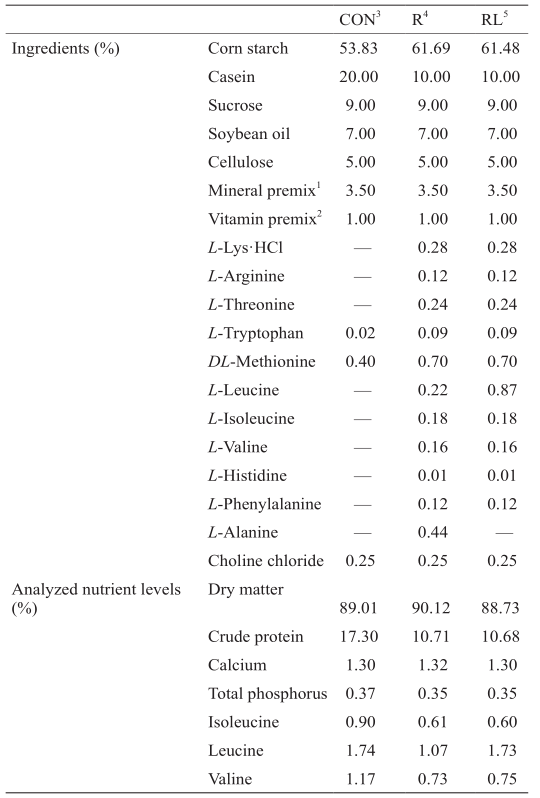
本试验选取36只12周龄、体重为(200 ±2.7) g的成年SD雄性大鼠(购于北京实验动物中心)。试验适应期为5 d,所有大鼠饲喂普通基础日粮(标准清洁级生长鼠料,AIN93-G日粮)。适应期结束后饲喂试验日粮,3种试验日粮分别含有20%酪蛋白的AIN93-G日粮(对照,20%C,CON)、10%酪蛋白 + 丙氨酸日粮(低蛋白日粮,10%C + Ala,R)、10%酪蛋白 + 亮氨酸日粮(10%C + Leu,RL),试验期共11 d。其中,CON组与RL组亮氨酸水平相同,R组与RL组含氮水平相同。日粮中其他营养水平均满足AIN-93G营养需要量[11],并参照此需要量分别在各处理组中添加适宜种类和数量的合成氨基酸。饲料加工由北京科澳协力饲料有限公司完成。3种日粮配方及营养水平见表1。
《表1》
表1 试验日粮的组成与营养水平(饲喂基础)
《2.2. 饲养管理》
2.2. 饲养管理
本试验在农业部饲料效价与安全监督检验测试中心(北京)鼠营养代谢室进行,试验鼠房为万级清洁实验室。大鼠饲喂在控温和控湿的房间内,温度为(20 ±2)℃,相对湿度为45% ± 10%。光照制度为昼夜光照交替12 h:12 h。大鼠单笼(20.0 cm × 17.5 cm × 19.5 cm)饲养,自由采食和饮水。
《2.3. 试验设计和样品采集》
2.3. 试验设计和样品采集
试验大鼠按体重相近的原则随机分为3个处理,每个处理12个重复,每个重复1只大鼠,分别饲喂3种试验日粮。适应期为5 d,正式试验期为11 d。于第10天晚上20:00将大鼠日粮移除,饥饿处理12 h,于第11天早晨08:00重饲试验日粮1 h。所有大鼠大剂量一次性腹腔注射150 μmol·(100 g–1 体重) L-苯丙氨酸,其中含有60 μmol·(100 g–1 体重) 的L-[ring- 2H5 ]苯丙氨酸(购买于美国Cambridge同位素实验室,L-苯丙氨酸用生理盐水溶解)[12]。腹腔注射在5~10 s内完成。30 min后,所有大鼠通过戊巴比妥钠麻醉,眼眶静脉采血于真空采血管(Becton Dickinson Vacutainer Systems,美国),4℃、2000 r·min–1 离心25 min,分离血浆,–80℃冻存备用。采血后,将大鼠颈部脱臼处死,迅速采集左侧腹脂、后肢的比目鱼肌、腓肠肌以及混合肌样品称重后,放入液氮中冷冻,之后于–80℃冻存用于骨骼肌蛋白质合成及相关指标的测定,而右侧肌肉样品放于预孵育液中孵育,用于测定骨骼肌蛋白质分解。
《2.4. 检测指标和方法》
2.4. 检测指标和方法
2.4.1. 日粮成分
日粮干物质、粗蛋白、总能、钙和磷的检测方法参照AOAC(2000)。
2.4.2. 日粮氨基酸含量的检测
试验日粮样品粉碎后分别在110℃下6 mol·L–1 盐酸水解24 h和0℃下过甲酸氧化16 h后再经盐酸水解24 h,用氨基酸自动分析仪(Hitachi L-8800 Amino Acid Ana-lyzer, Tokyo, Japan)测定15种氨基酸和含硫氨基酸含量;粉碎后的日粮用4 mol·L–1 氢氧化钠在110℃下水解22 h后,使用高效液相色谱仪(Hitachi, Tokyo, Japan)测定色氨酸含量[13]。
2.4.3. 生长性能
在试验第1天和第11天早晨分别对大鼠进行称重,并准确记录试验期大鼠采食量,计算大鼠在整个试验期的增重、平均日增重和饲料增重比。
2.4.4. 骨骼肌和腹脂重量的测定
试验结束称重后,每只大鼠注射含有L-[ring- 2H5 ]苯丙氨酸的氨基酸溶液,30 min后,采血并颈部脱臼处死,采集腹脂、后肢的比目鱼肌和腓肠肌样品称重。
2.4.5. 血清中游离氨基酸含量的测定
移取0.5 mL血清样品于5 mL离心管中,加入1.5 mL的10%磺基水杨酸和0.175 mL锂稀释液,振荡混匀后,冰上静置20 min,取上清液2 mL于4℃ 50 000 r·min–1 离心30 min,取上清液用0.1 µm滤膜过滤,装入上机小瓶中,上机,根据茚三酮柱后衍生测定血清游离氨基酸浓度。
2.4.6. 骨骼肌蛋白质合成的测定
骨骼肌的蛋白质合成测定具体过程如下[12,14]:用研钵将肌肉样品磨碎后,用2 mL 2 mol·L–1 的三氯乙酸溶液将样品(0.08 g)溶解,在冰上匀浆,之后4℃2000 r·min–1 离心20 min。将上清液(含有游离态苯丙氨酸)倒出,过阳离子交换树脂柱。沉淀部分(含有结合态苯丙氨酸)则用2 mL 2 mol·L –1 的三氯乙酸溶液洗涤3次,再用3 mL 6 mol·L –1 的盐酸溶液在110℃水解24 h,冷却、过滤和蒸干后,用超纯水溶解,过阳离子交换树脂柱。阳离子交换树脂柱先用1 mol·L –1 盐酸洗涤,然后用超纯水洗至中性,游离和结合氨基酸用4 mL4 mol·L –1 的氨水溶液洗脱,并在充氮环境中将收集的样品溶液蒸干[15]。样品中的游离态和结合态L-[ring- 2H5 ]苯丙氨酸经七氟丁酸酐衍生后,使用气相色谱-质谱仪(gas chromatography-mass spectrometry,GC-MS)测定其中游离态的苯丙氨酸检测电子质荷比为91和96时的丰度,而结合态的苯丙氨酸检测电子质荷比为148和153时的丰度,根据标准曲线,换算为摩尔分数(mol%)表示的丰度。
蛋白质相对合成率(FSR)计算方法参照Garlick等(1983),公式如下:
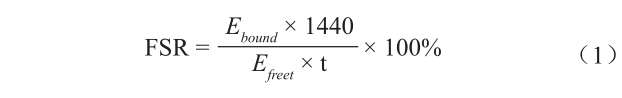
式中,FSR为蛋白质的相对合成率,定义为1 d内蛋白质更新的百分率,%·d–1 ;Ebound 为结合态L-[ring- 2H5 ]苯丙氨酸的同位素丰度,%;1440表示1 d中的时间,min·d–1 ;E freet 为在时间t时,游离态L-[ring- 2H5 ]苯丙氨酸的同位素丰度,%;t为从腹腔同位素注射后到处死取样的时间,在本试验中为30 min。
2.4.7. 肌 肉 中 磷 酸 化 4EBP1 和 S6K1 的 蛋 白 质 印 迹(Western blotting)测定
取出肌肉样品,利用RIPA裂解液裂解,加入蛋白酶抑制剂(南京建成生物工程研究所,江苏,中国)提取蛋白质,用BCA蛋白质测定试剂盒(Thermo Scientific,Rockford,IL,美国)测定蛋白质浓度。每孔上样相同的总蛋白质进行十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE),湿转法于100 V恒压冰上转膜1 h左右,分离胶中蛋白质被转至PVDF膜(上海康成生物公司有限公司,中国)。将PVDF膜浸没于5%的脱脂奶粉中,室温封闭1 h。封闭后的PVDF膜分别放在4EBP1一抗和S6K1一抗(Cell Signaling Technology,美国)溶液(按抗体说明书稀释)中4℃轻摇孵育过夜。用1×封闭缓冲液洗膜5次,每次5 min。用5%脱脂奶粉-TBST稀释二抗(辣根过氧化物酶HRP标记),室温轻摇孵育1 h。用1×封闭缓冲液洗膜5次,每次5 min。将化学发光试剂盒中两种试剂按比例混合为反应液,将膜置于反应液中室温孵育5 min。去除过量的溶液,将膜夹在两塑料薄膜之间,以X光胶片曝光,待胶片显影定影后,漂洗晾干,将胶片扫描保存为计算机文件,并用AlphaImager2200(Alpha Innotech,CA,USA)分析软件将图片上每个特异条带的灰度值数字化。内参蛋白选择β-肌动蛋白(β-actin)。
《2.5. 统计分析》
2.5. 统计分析
本试验测定结果以每只大鼠为统计单位,数据采用SAS 8.1版软件中单因素方差分析进行,差异显著时采用Duncan’s多重比较。当P < 0.05时确定为差异显著。
《3. 结果》
3. 结果
《3.1. 对成年大鼠生长性能的影响》
3.1. 对成年大鼠生长性能的影响
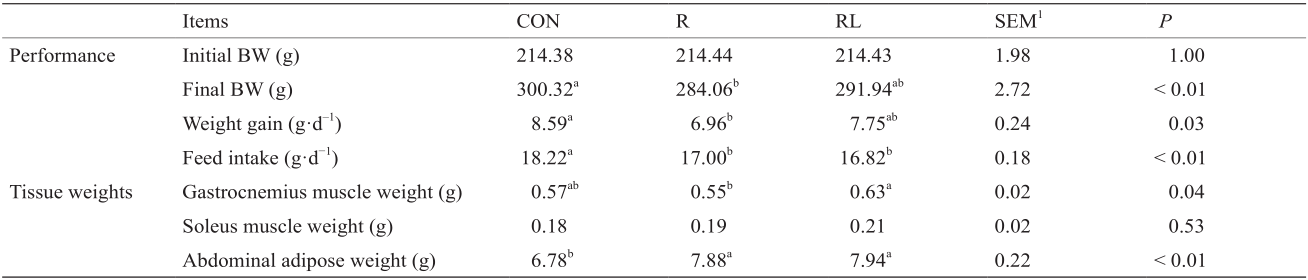
低蛋白日粮添加亮氨酸对成年大鼠生长性能的影响见表2。3个处理间初始体重无显著差异。试验日粮处理后,与对照组相比,10%C + Ala(R)组大鼠的最终体重和增加体重显著降低(P < 0.05),而10%C + Leu(RL)组差异不显著(P > 0.05)。与两个低蛋白日粮组相比,CON组的采食量显著升高(P < 0.05)。与R组相比,RL组可以显著增加腓肠肌重量(P < 0.05);而三个处理间的比目鱼肌重量无显著差异。与R组和RL组相比,CON组显著降低腹脂重量(P < 0.05)。
《表2 》
表2 低蛋白日粮添加亮氨酸对成年大鼠生长性能的影响
《3.2. 对血清氨基酸浓度的影响》
3.2. 对血清氨基酸浓度的影响
与CON组相比,R和RL组会显著升高血清中必需氨基酸浓度,如缬氨酸、精氨酸、赖氨酸、蛋氨酸、苏氨酸、组氨酸和色氨酸(P < 0.05,表3),同时显著升高非必需氨酸浓度,如丝氨酸、甘氨酸和丙氨酸(P <0.05)。与R组相比,RL日粮会显著升高亮氨酸、组氨酸和甘氨酸含量(P < 0.05),显著降低异亮氨酸含量(P< 0.05)。与CON组相比,RL日粮还显著增加亮氨酸含量(P < 0.05)。CON与R两处理在异亮氨酸和亮氨酸无显著差异。
《表3》
表3 低蛋白日粮添加亮氨酸对成年大鼠血清游离氨基酸浓度的影响(µmol· –1 )
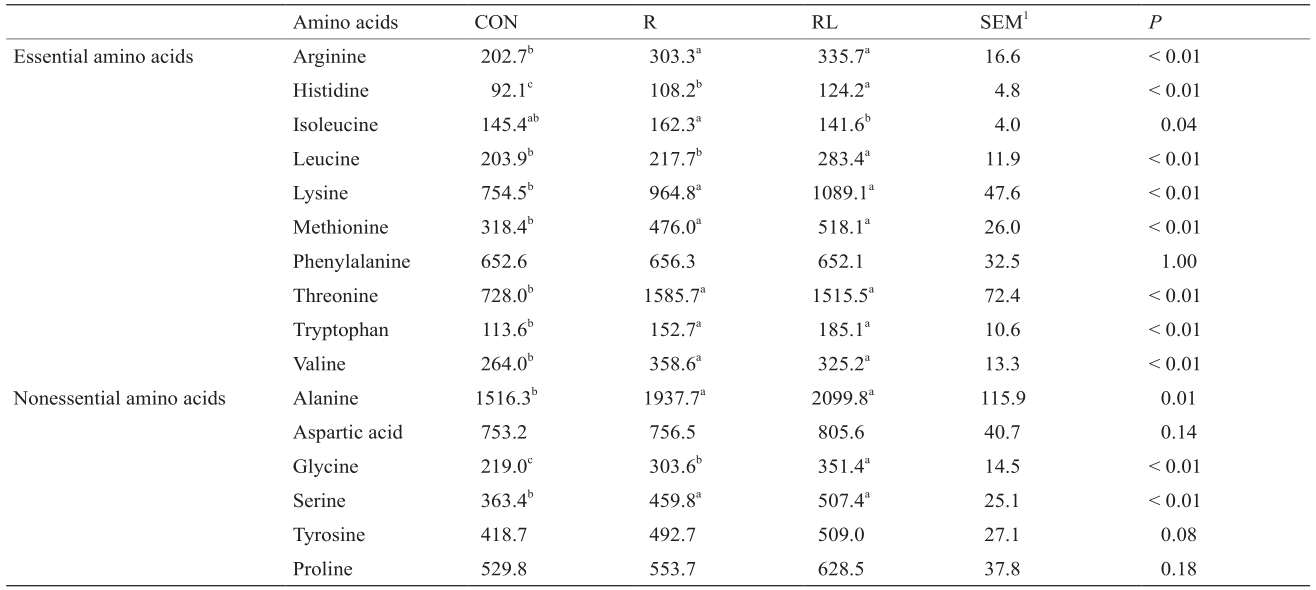
1SEM: standard error of the mean; the letters a,b,c indicate that values with different superscripts within a row are significantly different at P < 0.05.
《3.3. 对蛋白质合成的影响》
3.3. 对蛋白质合成的影响
与R和RL相比,CON日粮显著增加腓肠肌和比目鱼肌蛋白质合成(P < 0.05,图1)。与R相比,RL日粮显著增加大鼠骨骼肌蛋白质合成(P < 0.05)。
《图1》
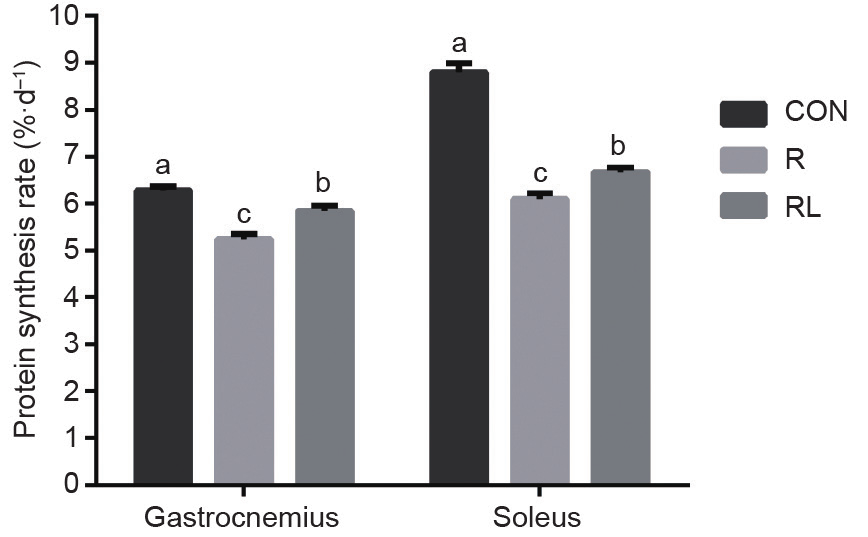
图 1. 低蛋白日粮添加亮氨酸对成年大鼠骨骼肌蛋白质合成的影响。CON:20%酪 蛋 白 日 粮;R:10%酪 蛋 白+0.44%丙 氨 酸 日 粮;RL:10%酪蛋白+0.87%亮氨酸日粮;a,b,c表示不同字母的值在P<0.05水平上差异显著。
《3.4. 对翻译因子磷酸化的影响》
3.4. 对翻译因子磷酸化的影响
CON组的4EBP1磷酸化显著高于R和RL(P < 0.05,图2)。S6K1磷酸化在RL组最高,显著高于R和CON组(P < 0.05,图3)。
《图2》
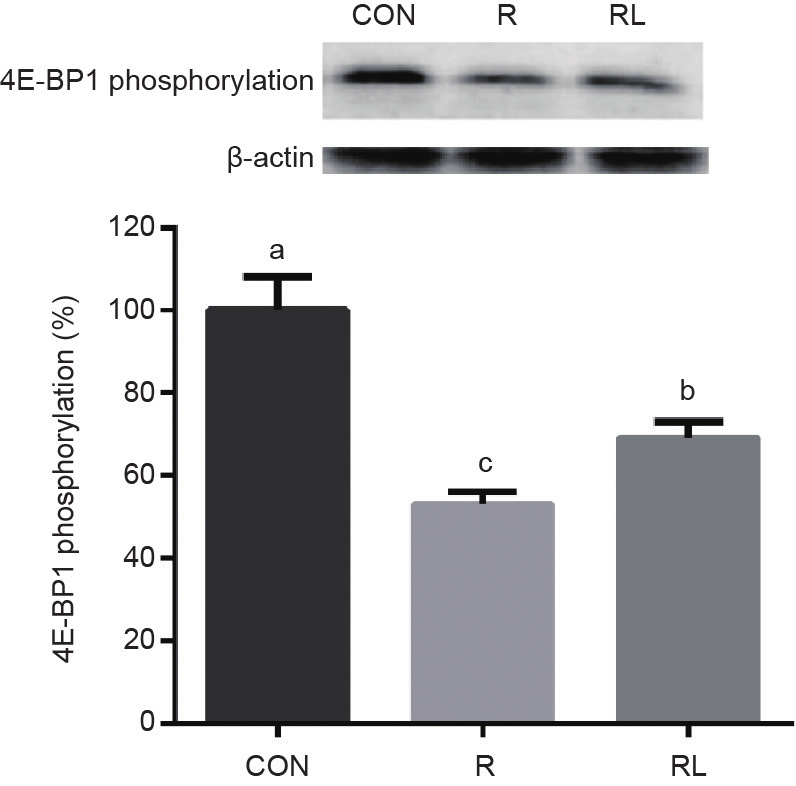
图2. 低蛋白日粮添加亮氨酸对成年大鼠骨骼肌中4E-BP1磷酸化的影响。 CON:20%酪蛋白日粮;R:10%酪蛋白+0.44%丙氨酸日粮;RL:10%酪蛋白+0.87%亮氨酸日粮;a,b,c表示不同字母的值在P<0.05水平上差异显著。
《图3》
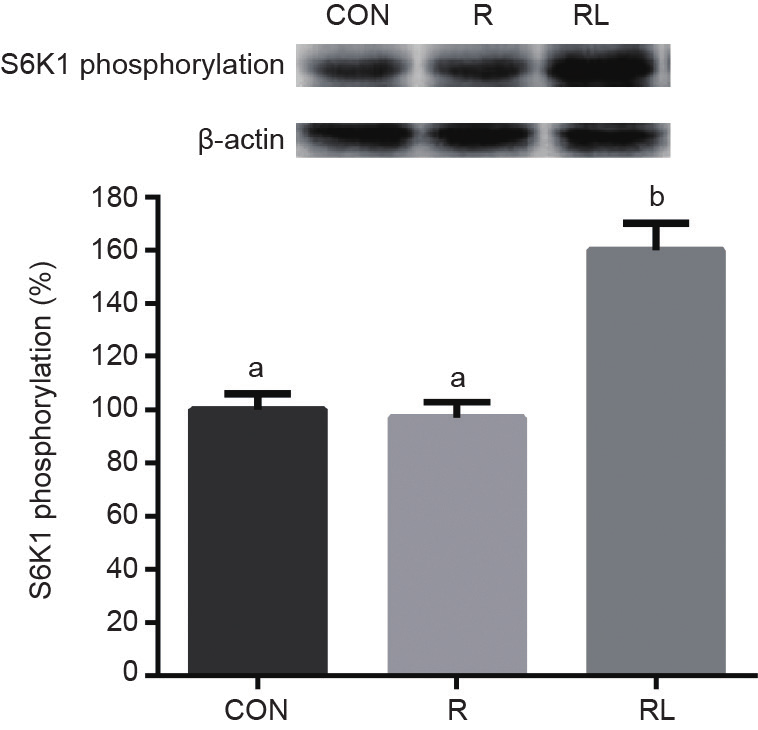
图3. 低蛋白日粮添加亮氨酸对成年大鼠骨骼肌中S6K1磷酸化的影响。CON:20%酪 蛋 白 日 粮;R:10%酪 蛋 白+0.44%丙 氨 酸 日 粮;RL:10%酪蛋白+0.87%亮氨酸日粮;a,b,c表示不同字母的值在P<0.05水平上差异显著。
《4. 讨论》
4. 讨论
临床中,一般建议有肾脏和肝脏疾病的患者减少蛋白质的摄入量。此外,由于饲料资源有限,为了节约饲料成本,低蛋白日粮在畜牧业中也得到了广泛的应用。但是研究表明,低蛋白摄入会降低人组织中蛋白质沉积[1]、减少新生仔猪的蛋白质合成[16]。已经报道短期添加亮氨酸到蛋白质严重缺乏的日粮中,可以减少大鼠体重损失和抑制骨骼肌的蛋白质降解[5]。但是长期添加亮氨酸对蛋白质合成的影响仍不明确,所以本试验探究
在成年大鼠的日粮中蛋白质严重缺乏时(蛋白质含量仅为大鼠需要量的60%),添加亮氨酸对肌肉蛋白质合成和生长性能的影响。
本试验结果表明,饲喂大鼠极低蛋白日粮(R组),即使按照大鼠需要量添加必需氨基酸,其体重增加仍显著低于CON组(P < 0.05)。而在低氮日粮中添加亮氨酸(RL组),使亮氨酸与CON组含量相同,则可以提高大鼠的体增重并与CON组无显著差异;同样RL组的腓肠肌重量与CON组差异不显著,并且显著重于R组(P< 0.05)。这表明亮氨酸添加到低蛋白日粮中,对大鼠的生长性能和肌肉质量具有促进作用。有研究表明,亮氨酸在新生仔猪体内被吸收后,其在血浆中的浓度升高两倍才能使蛋白质合成速度最大[8]。在本文中,与R组相比,RL组添加亮氨酸可以显著地提高血清中亮氨酸含量(P < 0.05),进而促进肌肉蛋白质合成(P < 0.05)。
尽管低蛋白日粮中添加亮氨酸增加了腓肠肌重量,并且3个组别的蛋白质合成速度存在显著性差异(图1),但比目鱼肌的重量在3个处理组之间无显著差异(表2)。这有可能是因为腓肠肌和比目鱼肌属于不同类型的肌肉,在腓肠肌中绝大部分是白肌肉纤维(类型2,快速收缩);而在比目鱼肌中绝大部分是红肌肉纤维(类型1,慢速收缩)。再者可能是因为大鼠腓肠肌的蛋白含量高于比目鱼肌[17],所以即使当比目鱼肌的蛋白质合成速度高于腓肠肌,添加亮氨酸只可能影响腓肠肌的重量而不会影响比目鱼肌。
此外,我们发现RL组的大鼠腹脂重量与R组差异不显著,但这两个低蛋白组腹脂的重量都显著重于CON组,这可能与低蛋白日粮的能量损失减少有关。研究表明,当日粮中蛋白质的含量减少时,可以减少多余氨基酸的去氨基反应及尿素在尿液中排泄,因此可以降低动物机体蛋白周转和热量产生,所以降低日粮蛋白含量可以增加组织中的可利用能量。并且在本研究中,为了降低日粮中的蛋白质含量,我们用玉米淀粉来替换日粮中的酪蛋白。相比酪蛋白,淀粉可以更高效地提供能量,有利于腹脂沉积。可以部分地解释本试验中低蛋白日粮增加脂肪沉积的结果。在本研究中添加亮氨酸到低蛋白日粮中并没有影响腹脂重量,但前人研究表明,给幼鼠饲喂添加亮氨酸日粮会增加腹脂25%[18],同样,Zeanandin等[19]也发现类似结果,他们认为亮氨酸组的腹脂增加与采食量增加或能量消耗减少有关。而Zhang等[20]则发现相反的结果,添加亮氨酸会增加能量消耗并减少腹脂。因此关于亮氨酸对腹脂的影响仍不清晰,需要进一步探究。
按照大鼠营养需要量[11],在低蛋白日粮R和RL组中添加了必需氨基酸。由于添加的晶体氨基酸能够快速地进入动物体内循环,所以能够被身体直接吸收利用。而在CON组日粮中只含有少量的晶体氨基酸(如色氨酸和蛋氨酸),绝大部分是以蛋白质形态存在。蛋白质首先被消化吸收,分解为游离的氨基酸才能被机体利用。这可以解释本试验中低蛋白日粮组的血清中大部分游离氨基酸的含量高于CON组。此外,与R组相比,CON组和RL组日粮的亮氨酸含量(接近1.74%)都显著高于R组日粮(1.07%),血清中游离的异亮氨酸浓度显著降低。这可能是由于亮氨酸对异亮氨酸代谢有拮抗作用[21]。这与前人的研究结果相一致[12,22,23],再次表明日粮中过多的亮氨酸可能降低血清中异亮氨酸浓度。关于亮氨酸诱导异亮氨酸含量变化可能的相关机理有两个:一方面可能与氨基酸转运载体影响氨基酸在体内的分布情况有关;另一方面可能与支链氨基酸的氧化有关[21]。
体内试验表明,仔猪的低蛋白日粮中添加亮氨酸可以促进骨骼肌的蛋白质合成[16],类似的结果在人的试验中也有报道[24]。体外试验同样表明,在培养基中添加0.5 mmol·L–1 的亮氨酸,可以促进受饥饿大鼠的比目鱼肌的蛋白质合成[25]。同样本试验中,与R组相比,RL组显著促进蛋白质合成(尽管低于CON组),缓解低蛋白对蛋白质合成的影响。这可能是由于血清中高浓度的亮氨酸能够促进蛋白质合成,这也可以部分解释RL组的蛋白质合成速度高于R组;但不能解释,CON组的蛋白质合成速度显著高于RL组,而亮氨酸浓度显著低于RL组。前人研究表明,成年人锻炼后长期补充亮氨酸对体内合成代谢的作用还取决于体内蛋白质合成所需的其他可利用底物的含量,因此,亮氨酸必须与其他的氨基酸同时使用来促进蛋白质的合成[26]。这也可以解释本试验中,CON组日粮中绝大部分必需氨基酸的含量高于低蛋白R和RL组,故CON组的蛋白质合成速率最高。
然而,本研究中RL和R组的绝大部分血清氨基酸含量高于CON组,这表明血清中氨基酸含量不太可能限制蛋白质合成。因此有必要考虑其他因素,尤其是在本试验中长期缺乏蛋白质的情况下影响蛋白质合成的因素。Kim等[27]研究表明,采食量影响蛋白质合成,只有采食足够多的蛋白质才能维持肌肉较高的蛋白质合成速率。此外,本试验中蛋白质合成速率与肌肉重量的结果不太一致。一方面是由于蛋白质合成速率并不等于肌肉重量,因为肌肉重量变化由蛋白质合成和降解的差值决定;另一方面与蛋白质合成速率测定方法有关,该测量方法适合于短时间处理,不太适合本研究中的长期试验[28]。其他研究也报道过类似结果[2,9]。
mTOR信号通路在促进细胞翻译起始和蛋白质合成起着重要作用[29,30],该信号通路有两个不同复合体,即mTORC1和mTORC2[31]。激活后的mTORC1对mRNA翻译起始因子S6K1和4EBP1磷酸化有促进作用。4EBP1经过磷酸化后会从eIF4E.4EBP1复合分子中释放出来,形成有活性的eIF4G.eIF4E复合物,然后与mRNA结合启动蛋白质翻译过程[29]。同样S6K1激活后可释放S6,S6参与到蛋白质翻译起始过程[29]。前人研究表明,亮氨酸可通过调节mTOR信号通路的下游因子S6K1和4EBP1磷酸化,来促进蛋白质合成[29,30]。最近研究表明,添加亮氨酸可促进正常仔猪和宫内发育受阻仔猪4EBP1和S6K1的磷酸化[32],新出生的仔猪也有相同结果[2]。而断奶仔猪长期饲喂蛋白质缺乏日粮,会抑制mTOR信号通路中4EBP1磷酸化,而不影响S6K1[9];这与本研究中图2和图3的结果一致。另外,本研究结果表明,添加亮氨酸可以缓解低蛋白处理产生的不利作用,但对4EBP1磷酸化促进作用仍显著低于CON组,这与蛋白质合成结果相一致。因此本试验验证了在蛋白质缺乏日粮中添加亮氨酸可通过S6K1磷酸化促进骨骼肌的蛋白质合成。但也有研究表明,通过饮水方式长期给幼年大鼠补充亮氨酸,可以促进肌肉蛋白质合成,但不影响S6K1和4EBP1的表达[33]。Vary[34]研究表明,给大鼠口服亮氨酸可促进蛋白质合成,主要通过4EBP1的磷酸化及减少4EBP1与EIF4E的结合,而不影响S6K1的活性。此外,Mao[12]等的研究表明,亮氨酸可通过上调瘦素受体的表达,来促进蛋白质合成。这些研究结果存在的差异,可能与试验动物品种和营养状况不同有关。
《5. 结论》
5. 结论
对于长期低蛋白日粮处理的大鼠而言,在日粮中添加亮氨酸可以缓解低蛋白日粮产生的不利作用,并通过mTOR信号通路促进蛋白质合成和提高骨骼肌重量。尽管本试验中,比目鱼肌中蛋白质的合成速度存在显著差异,但亮氨酸可以增加腓肠肌重量而不影响比目鱼肌。这可能与比目鱼肌和腓肠肌属于不同类型肌肉及蛋白质合成速率的测定方法有关。因此,需要进一步研究添加亮氨酸对不同类型的心脏和肌肉蛋白合成的影响。
《致谢》
致谢
本研究得到了北京食品营养与人类健康高精尖创新中心的资金支持。
《Compliance with ethics guidelines》
Compliance with ethics guidelines
Bo Zhang, Licui Chu, Hong Liu, Chunyuan Xie, Shiyan Qiao, and Xiangfang Zeng declare that they have no conflict of interest or financial conflicts to disclose.











 京公网安备 11010502051620号
京公网安备 11010502051620号




