
2022, Volume 16, Issue 9
Engineering >> 2022, Volume 16, Issue 9 doi: 10.1016/j.eng.2021.03.028
Genetic and Functional Differences of Escherichia coli Strains from Colorectal Cancer Mucosal Tissues
a State Key Laboratory of Pathogen and Biosecurity, Beijing Institute of Microbiology and Epidemiology, Beijing 100071, China
b Beijing Shijitan Hospital, Capital Medical University, Beijing 100038, China
c Guangdong Provincial Key Laboratory of Gastroenterology, Institute of Gastroenterology of Guangdong Province, &Department of Gastroenterology, Nanfang Hospital, Southern Medical University, Guangzhou 510515, China
d Shenzhen Center for Disease Control and Prevention, Shenzhen 518055, China
# These authors contributed equally to this work.
Next Previous
Abstract
Colorectal cancer (CRC) is the third leading cancer globally. Metagenomics has been widely used to analyze the association between the gut microbiota and CRC based on bacterial genus- or species-level comparisons, providing evidence of dysbiosis in CRC development. However, this kind of analysis is unable to provide strain-level information for understanding the individual role of a species in CRC. Here, we used culturomics to isolate CRC mucosal samples and selected 158 Escherichia coli strains to reveal their differences in both genomics and functions by means of phylogenetic analysis and inflammatory induction based on cell and animal experiments. Through genomic comparison, these strains were divided into five phylogroups. The representative strains of each phylogroup significantly induced different levels of cytokine secretion by human leukemic monocyte (THP-1 cell)-based Transwell and animal experiments. Further bioinformatic analysis revealed different profiles of single-nucleotide polymorphisms, genes, and metabolic pathways in the different phylogroups, which can improve the current understanding of the phenotypic differences between these strains. The strain differences revealed in both genomics and functions indicate that the microbiota’s function at the strain level should be investigated in order to understand the interacting mechanisms between hosts and gut bacteria.
Keywords
Colorectal cancer ; Gut microbiota ; Culturomics ; Escherichia coli ; Strain level
SupplementaryMaterials
Figures

Fig. 1
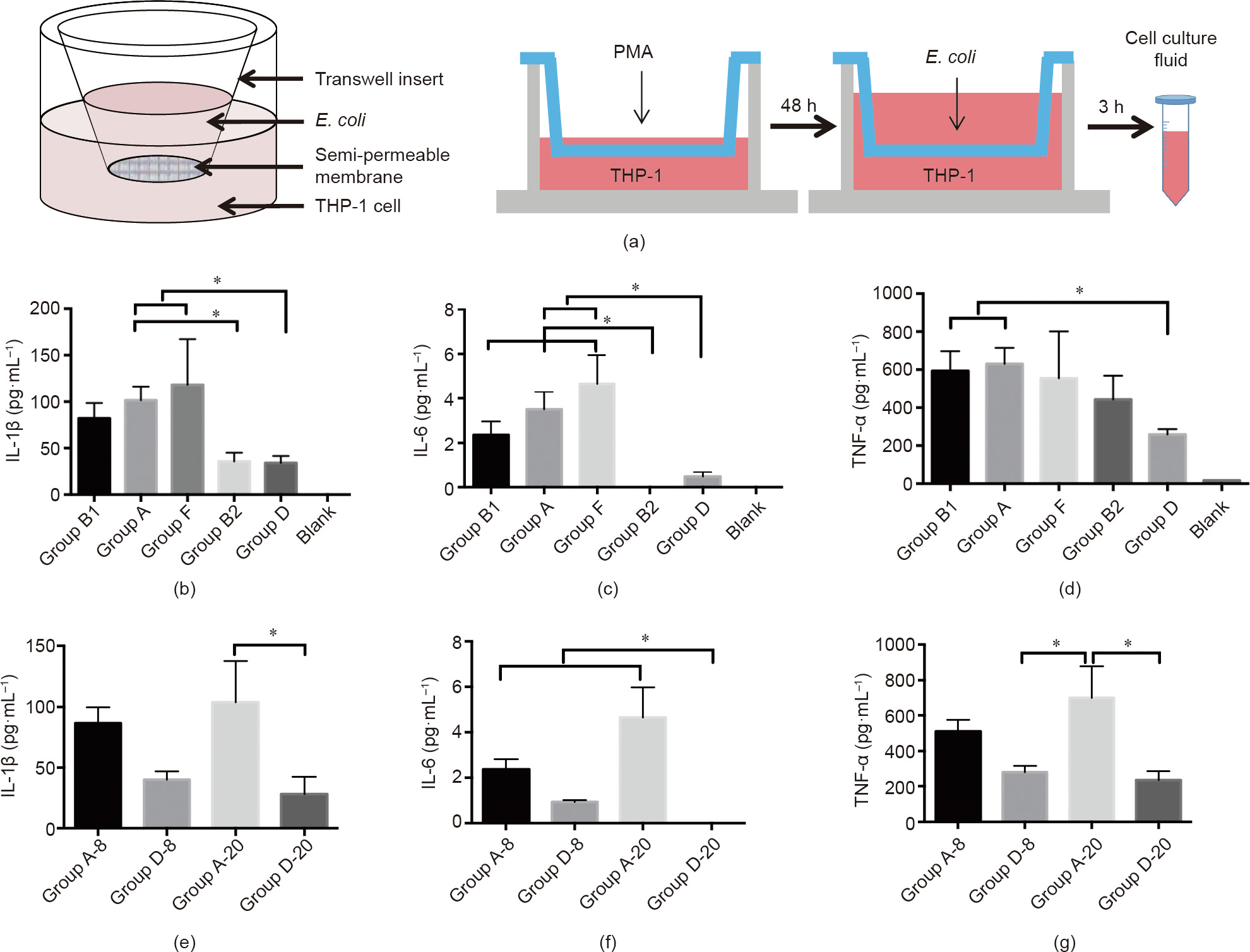
Fig. 2

Fig. 3

Fig. 4

Fig. 5
References
[ 1 ] Louis P, Hold GL, Flint HJ. The gut microbiota, bacterial metabolites and colorectal cancer. Nat Rev Microbiol 2014;12(10):661–72. link1
[ 2 ] Peters BA, Dominianni C, Shapiro JA, Church TR, Wu J, Miller G, et al. The gut microbiota in conventional and serrated precursors of colorectal cancer. Microbiome 2016;4(1):69. link1
[ 3 ] Sun J, Kato I. Gut microbiota, inflammation and colorectal cancer. Genes Dis 2016;3(2):130–43. link1
[ 4 ] Yu J, Feng Q, Wong SH, Zhang D, Liang QY, Qin Y, et al. Metagenomic analysis of faecal microbiome as a tool towards targeted non-invasive biomarkers for colorectal cancer. Gut 2017;66(1):70–8. link1
[ 5 ] Yang Y, Weng W, Peng J, Hong L, Yang L, Toiyama Y, et al. Fusobacterium nucleatum increases proliferation of colorectal cancer cells and tumor development in mice by activating Toll-like receptor 4 signaling to nuclear factor-jB, and up-regulating expression of microRNA-21. Gastroenterology 2017;152(4):851–66.e24. link1
[ 6 ] Long X, Wong CC, Tong L, Chu ESH, Ho Szeto C, Go MYY, et al. Peptostreptococcus anaerobius promotes colorectal carcinogenesis and modulates tumour immunity. Nat Microbiol 2019;4(12):2319–30. link1
[ 7 ] Barboza M, Sela DA, Pirim C, LoCascio RG, Freeman SL, German JB, et al. Glycoprofiling bifidobacterial consumption of galacto-oligosaccharides by mass spectrometry reveals strain-specific, preferential consumption of glycans. Appl Environ Microbiol 2009;75(23):7319–25. link1
[ 8 ] Lopez-Siles M, Khan TM, Duncan SH, Harmsen HJM, Garcia-Gil LJ, Flint HJ. Cultured representatives of two major phylogroups of human colonic Faecalibacterium prausnitzii can utilize pectin, uronic acids, and host-derived substrates for growth. Appl Environ Microbiol 2012;78(2):420–8. link1
[ 9 ] Wilson ID, Nicholson JK. Gut microbiome interactions with drug metabolism, efficacy, and toxicity. Transl Res 2017;179:204–22. link1
[10] Lloyd-Price J, Mahurkar A, Rahnavard G, Crabtree J, Orvis J, Hall AB, et al. Strains, functions and dynamics in the expanded Human Microbiome Project. Nature 2017;550(7674):61–6. link1
[11] Scholz M, Ward DV, Pasolli E, Tolio T, Zolfo M, Asnicar F, et al. Strain-level microbial epidemiology and population genomics from shotgun metagenomics. Nat Methods 2016;13(5):435–8. link1
[12] Lagier JC, Hugon P, Khelaifia S, Fournier PE, La Scola B, Raoult D. The rebirth of culture in microbiology through the example of culturomics to study human gut microbiota. Clin Microbiol Rev 2015;28(1):237–64. link1
[13] Sorbara MT, Littmann ER, Fontana E, Moody TU, Kohout CE, Gjonbalaj M, et al. Functional and genomic variation between human-derived isolates of Lachnospiraceae reveals inter- and intra-species diversity. Cell Host Microbe 2020;28(1):134–46.e4. link1
[14] Allali I, Delgado S, Marron PI, Astudillo A, Yeh JJ, Ghazal H, et al. Gut microbiome compositional and functional differences between tumor and non-tumor adjacent tissues from cohorts from the US and Spain. Gut Microbes 2015;6(3):161–72. link1
[15] Nakatsu G, Li X, Zhou H, Sheng J, Wong SH, Wu WKK, et al. Gut mucosal microbiome across stages of colorectal carcinogenesis. Nat Commun 2015;6 (1):8727. link1
[16] Ingala MR, Simmons NB, Wultsch C, Krampis K, Speer KA, Perkins SL. Comparing microbiome sampling methods in a wild mammal: fecal and intestinal samples record different signals of host ecology, evolution. Front Microbiol 2018;9:803. link1
[17] Didelot X, Falush D. Inference of bacterial microevolution using multilocus sequence data. Genetics 2007;175(3):1251–66. link1
[18] Jaureguy F, Landraud L, Passet V, Diancourt L, Frapy E, Guigon G, et al. Phylogenetic and genomic diversity of human bacteremic Escherichia coli strains. BMC Genomics 2008;9(1):560. link1
[19] Schuijt TJ, Lankelma JM, Scicluna BP, de Sousa e Melo F, Roelofs JJTH, de Boer JD, et al. The gut microbiota plays a protective role in the host defence against pneumococcal pneumonia. Gut 2016;65(4):575–83. link1
[20] Tenaillon O, Skurnik D, Picard B, Denamur E. The population genetics of commensal Escherichia coli. Nat Rev Microbiol 2010;8(3):207–17. link1
[21] Caugant DA, Levin BR, Selander RK. Genetic diversity and temporal variation in the E. coli population of a human host. Genetics 1981;98 (3):467–90. link1
[22] Bian Y, Du Y, Wang R, Chen N, Du X, Wang Y, et al. A comparative study of HAMSCs/HBMSCs transwell and mixed coculture systems. IUBMB Life 2019;71 (7):1048–55. link1
[23] Croxen MA, Law RJ, Scholz R, Keeney KM, Wlodarska M, Finlay BB. Recent advances in understanding enteric pathogenic Escherichia coli. Clin Microbiol Rev 2013;26(4):822–80. link1
[24] Micenková L, Bosák J, Vrba M, Ševcˇíková A, Šmajs D. Human extraintestinal pathogenic Escherichia coli strains differ in prevalence of virulence factors, phylogroups, and bacteriocin determinants. BMC Microbiol 2016;16(1):218. link1
[25] Lindstedt BA, Finton MD, Porcellato D, Brandal LT. High frequency of hybrid Escherichia coli strains with combined Intestinal Pathogenic Escherichia coli (IPEC) and Extraintestinal Pathogenic Escherichia coli (ExPEC) virulence factors isolated from human faecal samples. BMC Infect Dis 2018;18(1):544. link1
[26] Raisch J, Buc E, Bonnet M, Sauvanet P, Vazeille E, de Vallée A, et al. Colon cancer-associated B2 Escherichia coli colonize gut mucosa and promote cell proliferation. World J Gastroenterol 2014;20(21):6560–72. link1
[27] Martinez-Medina M, Garcia-Gil LJ. Escherichia coli in chronic inflammatory bowel diseases: an update on adherent invasive Escherichia coli pathogenicity. World J Gastrointest Pathophysiol 2014;5(3):213–27. link1
[28] Camprubí-Font C, Ewers C, Lopez-Siles M, Martinez-Medina M. Genetic and phenotypic features to screen for putative adherent-invasive Escherichia coli. Front Microbiol 2019;10:108. link1
[29] Prorok-Hamon M, Friswell MK, Alswied A, Roberts CL, Song F, Flanagan PK, et al. Colonic mucosa-associated diffusely adherent afaC+ Escherichia coli expressing lpfA and pks are increased in inflammatory bowel disease and colon cancer. Gut 2014;63(5):761–70. link1
[30] Suresh A, Ranjan A, Jadhav S, Hussain A, Shaik S, Alam M, et al. Molecular genetic and functional analysis of pks-harboring, extra-intestinal pathogenic Escherichia coli from India. Front Microbiol 2018;9:2631. link1
[31] Nougayrède JP, Homburg S, Taieb F, Boury M, Brzuszkiewicz E, Gottschalk G, et al. Escherichia coli induces DNA double-strand breaks in eukaryotic cells. Science 2006;313(5788):848–51. link1
[32] Gur C, Ibrahim Y, Isaacson B, Yamin R, Abed J, Gamliel M, et al. Binding of the Fap2 protein of Fusobacterium nucleatum to human inhibitory receptor TIGIT protects tumors from immune cell attack. Immunity 2015;42(2):344–55. link1
[33] Kantele A, Lääveri T, Mero S, Häkkinen IMK, Kirveskari J, Johnston BD, et al. Despite predominance of uropathogenic/extraintestinal pathotypes among travel-acquired extended-spectrum b-lactamase-producing Escherichia coli, the most commonly associated clinical manifestation is travelers’ diarrhea. Clin Infect Dis 2020;70(2):210–8. link1
[34] Terlizzi ME, Gribaudo G, Maffei ME. UroPathogenic Escherichia coli (UPEC) infections: virulence factors, bladder responses, antibiotic, and non-antibiotic antimicrobial strategies. Front Microbiol 2017;8:1566. link1
[35] Wijetunge DSS, Gongati S, DebRoy C, Kim KS, Couraud PO, Romero IA, et al. Characterizing the pathotype of neonatal meningitis causing Escherichia coli (NMEC). BMC Microbiol 2015;15(1):211. link1
[36] Tyakht AV, Manolov AI, Kanygina AV, Ischenko DS, Kovarsky BA, Popenko AS, et al. Genetic diversity of Escherichia coli in gut microbiota of patients with Crohn’s disease discovered using metagenomic and genomic analyses. BMC Genomics 2018;19(1):968. link1
[37] Chang PH, Pan YP, Fan CW, Tseng WK, Huang JS, Wu TH, et al. Pretreatment serum interleukin-1b, interleukin-6, and tumor necrosis factor-a levels predict the progression of colorectal cancer. Cancer Med 2016;5(3):426–33. link1
[38] Ray AL, Berggren KL, Restrepo Cruz S, Gan GN, Beswick EJ. Inhibition of MK2 suppresses IL-1b, IL-6, and TNF-a-dependent colorectal cancer growth. Int J Cancer 2018;142(8):1702–11. link1
[39] Spurbeck RR, Dinh PC, Walk ST, Stapleton AE, Hooton TM, Nolan LK, et al. Escherichia coli isolates that carry vat, fyuA, chuA, and yfcV efficiently colonize the urinary tract. Infect Immun 2012;80(12):4115–22. link1
[40] Rolhion N, Barnich N, Claret L, Darfeuille-Michaud A. Strong decrease in invasive ability and outer membrane vesicle release in Crohn’s diseaseassociated adherent-invasive Escherichia coli strain LF82 with the yfgL gene deleted. J Bacteriol 2005;187(7):2286–96. link1
[41] Alp D, Kuleasan H, Korkut Altıntas A. The importance of the S-layer on the adhesion and aggregation ability of lactic acid bacteria. Mol Biol Rep 2020;47 (5):3449–57. link1
[42] Tapader R, Bose D, Basu P, Mondal M, Mondal A, Chatterjee NS, et al. Role in proinflammatory response of YghJ, a secreted metalloprotease from neonatal septicemic Escherichia coli. Int J Med Microbiol 2016;306 (7):554–65. link1
[43] Tapader R, Bose D, Dutta P, Das S, Pal A, Raffatellu M. SslE (YghJ), a cellassociated and secreted lipoprotein of neonatal septicemic Escherichia coli, induces Toll-like receptor 2-dependent macrophage activation and proinflammation through NF-jB and MAP kinase signaling. Infect Immun 2018;86(9):e00399-18. link1
[44] Ahern P, Faith J, Gordon J. Mining the human gut microbiota for effector strains that shape the immune system. Immunity 2014;40(6):815–23. link1
[45] Bron PA, van Baarlen P, Kleerebezem M. Emerging molecular insights into the interaction between probiotics and the host intestinal mucosa. Nat Rev Microbiol 2012;10(1):66–78. link1











 京公网安备 11010502051620号
京公网安备 11010502051620号




