

《1.Introduction》
1.Introduction
Conventional swine diets contain substantial amounts of cereal grains (e.g., corn and wheat) and protein supplements such as soybean meal to provide pigs with the energy and nutrients they require. However, recent trends in the demand and supply of these conventional feedstuffs require swine producers around the world to seek low-cost alternatives such as cereal co-products from the biofuel and milling industries to feed their pigs, in order to reduce feed costs [1]. The majority of these co-products have a high energy and nutrient content but are fibrous in nature. When fibrous co-products are incorporated into pig diets, the carbohydrate composition inevitably changes from a high-starch diet toward a diet containing less starch and more non-starch polysaccharides, which are the major component of dietary fiber.
In general, diets that are rich in dietary fiber have a lower nutritive value for monogastric animals, including pigs, because these animals’ digestive enzymes are not suited to degrading non-starch polysaccharides [2]. The ingestion of high-fiber diets also has the potential to adversely affect energy and nutrient utilization and consequently result in lower pig performance [3–6]. Therefore, only a minimal level of fiber is typically included in diets fed to swine. However, dietary fiber has received a considerable amount of attention in recent years because some fiber components have beneficial effects on pigs when fermented in the intestine [7,8], and can also affect satiety and animal behavior [9,10].
This review discusses dietary fiber, its effects in pig nutrition, and the mechanisms involved in its utilization. Furthermore, the effects of dietary fiber on pig gut health and sow welfare are discussed. Finally, areas that require further research to expand our knowledge on fiber and some of the strategies used to improve fiber utilization in pigs are outlined.
《2.Dietary fiber》
2.Dietary fiber
《2.1. Definition and classification》
2.1. Definition and classification
Plant carbohydrates can be classified as sugars, disaccharides, oligosaccharides, or polysaccharides (i.e., starch and non-starch polysaccharides). Sugars, oligosaccharides, and starch are found in the interior of the plant cell, whereas non-starch polysaccharides, together with lignin, are the main constituents of the plant cell walls, and are called dietary fiber. The term “dietary fiber” has several definitions; however, all these definitions have limitations because plant cell wall components are variable and complex in their chemical and physical composition and in their metabolic effects. The initial definition of dietary fiber as “the sum of lignin and cell wall polysaccharides that are resistant to enzymatic hydrolysis in the digestive system of man” was coined by Trowell et al. [11] in relation to human medicine. However, this definition of dietary fiber is also applicable to other monogastric animals, such as pigs [12]. The Codex Alimentarius Commission [13] finalized the definition of dietary fiber as “carbohydrate polymers with 10 or more monomeric units, which are not hydrolyzed by the endogenous enzymes in the small intestine of humans.” Dietary fiber also includes any polysaccharides that reach the hindgut, such as resistant starch and oligosaccharides, which constitute plant cell contents and include fructo-oligosaccharides. The main constituents of the plant cell wall polysaccharides are cellulose, hemicellulose, and pectin.
Cellulose is a linear polymer of glucose units with β-(1→4) linkages, whereas pectin consists mainly of glucuronic acid units joined in chains by α-(1→4) glycosidic linkages. The most abundant organic substrate on earth, cellulose forms the main structural component of plant cell walls. Hemicelluloses are a complex matrix of polysaccharides that include xylose, arabinose, galactose, mannose, glucuronic acid, and β-glucans. Lignin is a phenolic polymer that anchors the cell wall polysaccharides and is not digested or fermented by porcine intestinal enzymes or bacteria, respectively [2,7,8,14].
Non-starch polysaccharides can be classified as insoluble or soluble based on their solubility in water or weak alkali [12]. Insoluble non-starch polysaccharides include cellulose and some hemicelluloses, and soluble non-starch polysaccharides include pectins, gums, and β-glucans. Soluble non-starch polysaccharides are more rapidly fermented in the gastrointestinal tract of the pig than insoluble non-starch polysaccharides [7,12]. For insoluble non-starch polysaccharides, little or no fermentation occurs in the upper gut; rather, fermentation remains low in the hindgut of pigs [15].
Dietary fiber (or non-starch polysaccharide fractions) is currently classified based on physicochemical properties in order to provide more information on its metabolic and physiological activities. The physicochemical properties of dietary fiber that are relevant to pig nutrition include viscosity, hydration, and fermentability. Viscosity describes the dissociation of non-starch polysaccharides in the gastrointestinal tract to form high molecular weight viscous aggregates. Soluble fibers such as β-glucans, gums, or pectins increase digesta viscosity when ingested by pigs. Fermentability describes the ability of non-starch polysaccharides to be fermented by the microflora harbored in the intestine. Soluble fiber is generally more fermentable than insoluble fiber. The hydration properties are swelling capacity, solubility, water-holding capacity, and water-binding capacity [16]. The water-holding capacity of a fiber affects its fermentability. Thus, when physicochemical parameters are incorporated into pig feed formulation, they may provide nutritionists with a better control of the fermentation process that takes place in the pig’s gut and could assist nutritionists in predicting the energy contribution and prebiotic effect of a diet or feedstuff.
《2.2. Analytical methods for characterizing dietary fiber》
2.2. Analytical methods for characterizing dietary fiber
Several methods exist for characterizing the dietary fiber component of feeds and feedstuffs; the choice of an analytical method depends on the aims of the investigator [12,17,18]. Based on how the fibrous remnants are isolated and measured, dietary fiber analytical methods are classified into three groups: chemical-gravimetric, enzymatic-gravimetric, and enzymatic-chemical methods. The numerous fiber analytical methods and the variability among these methods and among the obtained results make it rather difficult to compare information from different studies [19]. However, methods that categorize dietary fiber into soluble and insoluble components seem to provide the most accurate interpretation of study results. In what follows, the analytical methods most commonly used for measuring dietary fiber are briefly discussed.
2.2.1. Crude fiber method
Crude fiber analysis is a chemical-gravimetric method that is part of the Weende proximate analysis used for feed ingredients [19]. It was introduced to differentiate between carbohydrate that is “available” and carbohydrate that is “unavailable” for digestion. The aim of crude fiber analysis is to mimic the digestive actions of the gastric and pancreatic secretions by boiling a feed with dilute acid followed by a dilute alkali solution [19]. The crude fiber analytical method is very robust and reproducible within and among laboratories; however, there is incomplete recovery of cellulose, hemicellulose, and lignin. Therefore, crude fiber is not considered to be an acceptable definition for dietary fiber and is not suitable for characterizing the fiber component in pig feed. However, many regulatory agencies use crude fiber for quality control purposes and for regulating the minimum fiber content allowed in a feed.
2.2.2. The detergent (Van Soest) methods
The detergent methods, which are chemical-gravimetric procedures, were developed by Van Soest in the 1960s. These methods employ detergents to progressively extract neutral detergent fiber (NDF), acid detergent fiber (ADF), and acid detergent lignin (ADL) [18,19]. The NDF procedure recovers the insoluble components of dietary fiber (i.e., cellulose, hemicellulose, and lignin) after digesting a feed or ingredient in a solution at a neutral pH. Thus, the nutritional advantages of the NDF procedure are that it is able to approximate the insoluble dietary fiber fraction of a feedstuff and that its results are reproducible [19]. The ADF procedure recovers mainly cellulose and lignin by digesting a diet or feedstuff in a solution at an acidic pH [18], whereas the ADL recovers lignin using sulfuric acid. A more accurate estimate of hemicellulose and cellulose is obtained when the NDF, ADF, and ADL are determined sequentially using the same sample [18].
The detergent procedures, although an improvement over the crude fiber method, do not recover soluble dietary fiber (e.g., pectins, gum, and β-glucans) [19]. Therefore, these procedures can underestimate the total dietary fiber, especially for starchy feeds or ingredient samples, although probably not for cereal co-products, which have a high insoluble fiber concentration.
2.2.3. Total dietary fiber method
The total dietary fiber method was introduced to overcome some of the pitfalls of the detergent procedure. The total dietary fiber method is important for feedstocks intended for monogastric animals, including pigs, with hindgut fermentation. This procedure employs enzymes to simulate the processes that take place within the digestive tract [19], and may separate dietary fiber into soluble and insoluble fiber. The carbohydrates included in the definition of total dietary fiber include celluloses, hemicelluloses, oligosaccharides, lignin, pectins, and gums. Thus, total dietary fiber differs from nonstarch polysaccharides because lignin is included in the definition. The total dietary fiber method recovers more of the fibrous components than the NDF method, although it does not recover oligosaccharides that are soluble in aqueous ethanol.
There are two major methods to this procedure: enzymatic-gravimetric and enzymatic-chemical methods. The enzymatic-gravimetric methods are used to measure both insoluble and soluble fractions of dietary fiber; the AOAC Official Method 985.29 is the final version of these methods [20]. This procedure includes treatment of the sample with enzymes to remove starch and protein, precipitation of soluble fiber with aqueous ethanol, isolation and weighing of the residue and, finally, correction for protein and ash in the residue [19,21]. The enzymatic-chemical methods also use enzymes to remove starch, followed by precipitation of the soluble non-starch polysaccharides with aqueous ethanol. Thereafter, the neutral sugars are quantified by gas-liquid chromatography or high-performance liquid chromatography, and uronic acid is quantified by colorimetry [19].
The Uppsala and Englyst methods are the two most common enzymatic-chemical methods available. The Uppsala method quantifies the non-starch polysaccharides portion as the sum of amylaseresistant polysaccharides, uronic acid, and lignin, whereas the Englyst method does not include lignin and resistant starch in the final value [22]. Although the total dietary fiber methods are an improvement over the detergent procedures, they are still time-consuming, laborious, and relatively expensive.
《3. Dietary fiber in pig nutrition》
3. Dietary fiber in pig nutrition
Starch and non-starch polysaccharides differ not only in chemical structure but also in the type of nutrients they supply. Starch digestion in the upper gut results in glucose, whereas non-starch polysaccharide degradation in the lower gut results in volatile fatty acids [16,23]. In addition, pigs do not secrete the digestive enzymes required to break down non-starch polysaccharides, whereas they do secrete the enzymes required to hydrolyze starch. Furthermore, starch and non-starch polysaccharides differ in terms of their effects on other nutrients in the digestion process and on the performance of pigs [16,23]. Several authors have reviewed aspects of high-fiber diets in pig nutrition. Taken together, the available information suggests that dietary fiber has both positive and negative effects on the pig.
Most of the information on dietary fiber in pig nutrition has been generated using purified fiber sources such as pectin, cellulose, and guar gum, along with partially identified fiber from sources such as wheat bran [12], as opposed to information generated using natural fibrous feedstuffs such as distillers dried grain with solubles. Natural fibrous feedstuffs are usually composed of more than one fiber type; that is, they are composed of both soluble and insoluble fiber. Therefore, the effect of natural fibrous feedstuffs on the digestive physiology of pigs may not necessarily be similar to that of purified fiber. In the following sections, fiber effects on growing pigs are discussed in the context of fiber that has been obtained from purified and natural sources.
《3.1. Effects of fiber on pig growth performance》
3.1. Effects of fiber on pig growth performance
The effect of high-fiber ingestion has generally been to reduce pig growth rate. However, the reports obtained have often been contradictory. For example, feeding growing pigs diets containing either 7% guar gum or 7% cellulose reduced growth rate and final body weight [4]. Increasing the inclusion rate of distillers dried grains with solubles from 0 to 20% in nursery pig diets [24] and from 0 to 30% in growing pig diets [5] linearly decreased pig body weight, although the diets were formulated to contain similar net energy and standard ileal digestibility amino acids. Results from some studies [25,26] suggest that the decline in pig growth rate that occurs when high-fiber diets are fed is generally limited to the growing period rather than the finishing period. Conversely, in some studies [27–29], high-fiber diets had no adverse effects on pig performance in the growing or finishing phases, compared with the control.
The abovementioned studies reported on various types of feeds that were termed “high-fiber” diets, which contained varying amounts of ingredients (i.e., purified and natural fiber sources). This may partly explain the varying responses to high-fiber diets in terms of pig growth, because different inclusion rates of high-fiber ingredients and fiber types will elicit different responses. Furthermore, the specific fiber components and analytical methods used to measure fiber differed in those studies. Thus, crude fiber, NDF, total dietary fiber, and non-starch polysaccharides cannot be used interchangeably as equal measures of dietary fiber when assessing the effects of high fiber ingestion on pig performance.
Diets that are bulky due to the addition of fibrous feedstuffs have been reported to depress feed intake in pigs due to gut fill [4,30,31]. In gestating sows, early satiety due to the intake of high-fiber diets prevents certain stereotypies and thus is paramount to sow welfare [10], whereas in growing pigs, reduced feed intake is detrimental to performance. However, in growing-finishing pigs, feed intake does not always decrease when the pigs are fed high-fiber diets. For example, feed intake did not decrease in growing-finishing pigs that were offered diets containing up to 30% cereal grain co-products from the ethanol industry in several experiments [5,6,32–34]. In pigs raised in a thermo-neutral environment, feed intake is typically influenced by the diets’ energy content rather than by the bulkiness of the diet [35]. The addition of fibrous feedstuffs to pig diets dilutes the dietary energy content. Therefore, a decrease in dietary energy content is typically associated with increased feed intake to compensate for the energy required to support the maintenance and growth of pigs. Nonetheless, a period of adaptation to a fibrous diet is required before growing pigs are able to consume enough of a high-fiber diet to compensate for the dietary energy dilution [35].
《3.2. Effects of fiber on nutrient digestibility》
3.2. Effects of fiber on nutrient digestibility
Pre-cecal and total tract nutrient digestibilities in pigs have been reported to be negatively affected by high-fiber diets. Several mechanisms have been put forward to explain the negative impact of high-fiber diets on nutrient digestion. In this context, soluble fiber has been reported to increase digesta viscosity and thereby slow down the diffusion of the substrate and enzymes in the porcine small intestine, which hampers nutrient digestion and absorption [3]. Indeed, it has been reported that soluble fiber from diets based on purified guar gum, sugar beet pulp, oats, wheat, and barley increased digesta viscosity, leading to a reduction in nutrient digestion [4,12,36]. On the other hand, insoluble fiber reduces digesta passage rate, thereby allowing less mixing time for digestive enzymes and dietary components [3]. For example, insoluble fiber from wheat bran, corn bran, and cellulose reduced digesta transit time in several studies, leading to decreased nutrient digestibility [4,10,37,38]. However, the fiber in distillers dried grains with solubles (predominately insoluble) had no effect on digesta transit time in the experiment by Urriola and Stein [39]. This suggests that the effect of insoluble fiber on digesta transit time may vary among studies, and probably depends on the presence of interfering factors that are likely to exist in fiber from natural feedstuffs. Nonetheless, insoluble non-starch polysaccharides have been reported to decrease digesta transit time in the hindgut, whereas digesta transit time in the small intestine is unaffected or reduced [3,37].
Diets that are rich in non-starch polysaccharides encapsulate nutrients and thereby hamper their accessibility to digestive enzymes for hydrolysis [3]. Ingestion of fibrous diets has been reported to reduce nutrient digestibility in pigs through increased endogenous intestinal nutrient losses [3,7,17,40,41]. In addition, ingestion of fibrous diets leads to increased thickness of the unstirred water layer adjacent to the intestinal mucosa, which can impair nutrient digestion and absorption [42].
3.2.1. Fiber effect on starch digestion
Although endogenous carbohydrate-degrading enzyme activities may be reduced by fiber ingestion [43], several studies [36,44,45] have reported that starch digestibility in pigs is not affected by dietary fiber content, and that dietary starch is almost completely digested at the end of the small intestine. The lack of a fiber effect on starch digestibility is suggested to be due to the relatively long length of the small intestine, which ensures that starch is digested efficiently before the end of the ileum [36]. However, Wenk [3] suggested that a fiber-rich diet may reduce digesta transit time and allow a greater amount of undigested starch to pass through the ileum. Indeed, Gao et al. [46] and Agyekum et al. [47] recently observed that the addition of 5% inulin or 30% distillers dried grains with solubles to a corn-soybean meal-based diet fed to growing-finishing pigs reduced ileal starch digestibility, probably due to increased ileal starch flow. Nonetheless, although overall starch digestibility may not be affected by dietary fiber, the rate of starch digestion and absorption decreases due to fiber ingestion [48,49].
The decrease in the rate of starch digestion has been attributed to the luminal viscosity and nutrient-encapsulating effect of intact cell wall polysaccharides [48]. Soluble fiber appears to have a greater impact on starch digestion and absorption than insoluble fiber [49,50]. The reduction in starch absorption that occurs when pigs are fed fiber-rich diets may also be due to the decreased dietary starch load, in comparison with low-fiber diets, which are typically rich in starch.
3.2.2. Fiber effect on protein digestion
The literature contains a plethora of information on the effect of fiber on protein digestion in pigs. In general terms, ingestion of a fibrous diet reduces both ileal and total tract protein digestibility. The reduction in ileal protein and amino acid digestibility due to fiber ingestion has been reported to be associated with increased endogenous and exogenous nitrogen and amino acid losses, whereas increased microflora (bacterial protein) excretion in the feces is primarily responsible for the reduction in total tract protein digestibility observed in pigs [3,17,51].
Endogenous ileal nitrogen and amino acid losses refer to the nitrogen or amino acids that are present in endogenously synthesized proteins and that have not been digested and reabsorbed before reaching the end of the ileum [17,51]. These losses can be classified into basal loss, which is the minimum quantity of nitrogen and amino acids that is inevitably lost in all diets, and specific losses, which are influenced by dietary ingredient composition [51].
Concerning diet-specific ileal endogenous nitrogen and amino acid losses, several studies have reported that ingestion of fibrous diets increases the secretion of digestive enzymes and juices to accommodate dietary fiber in the digestive tract. For example, Langlois et al. [52] reported that a diet containing 40% wheat bran increased pancreatic juice and enzyme secretion in growing pigs. Increases in bile juice, mucin, and sloughed epithelial cells have also been reported to be due to fiber ingestion in pigs [17,41,53]. Digestive juices, mucin, and sloughed epithelial cells are rich in nitrogen and amino acids. Therefore, when fiber ingestion causes an increase in their production and a decrease in their reabsorption before the end of the ileum, the result is decreased ileal nitrogen and amino acid digestibility. Dietary fiber can also adsorb amino acid and peptides, thereby withholding them from digestion in the small intestine [17] and leading to decreased ileal nitrogen and amino acid digestibility.
3.2.3. Fiber effect on lipid digestion
The effect of fiber on lipid digestibility has been investigated extensively in pigs. Shi and Noblet [54] reported that ingestion of high-fiber diets decreased lipid digestibility in pigs. Similarly, feeding growing pigs cereal-based diets containing 20% or 40% wheat bran decreased lipid digestibility [37]. Dietary fiber depresses lipid digestibility through several mechanisms. For example, dietary fiber can increase digesta passage rate and thereby limit the time required by digestive enzymes to digest dietary lipids [55,56]. Alternatively, dietary fiber can bind with bile salts in the duodenum and render bile salts unavailable for the emulsification of dietary lipids. This in turn requires the synthesis of additional bile salts from blood cholesterol and thereby lowers blood cholesterol [55,56].
Reduction in lipid digestibility has been reported to be dependent on the type of fiber. Soluble fiber increases digesta viscosity and thereby prevents substrate-enzyme interaction and decreases lipid digestion, whereas insoluble fiber reduces digesta transit time in the digestive tract and thus increases lipid excretion in the feces [57]. However, not all studies have reported a negative effect of fiber-rich diets on lipid digestion. For example, the addition of wheat bran, corn bran, soybean hulls, and sugar beet pulp to increase the fiber content of diets fed to pigs did not affect lipid digestibility relative to the control diet [38]. Kil et al. [58] also observed that the dietary inclusion of purified NDF (from Solka-Floc) did not affect lipid digestibility relative to the control. Conversely, Högberg and Lindberg [45] reported that feeding growing pigs diets that contained the brans from oats, wheat, and rye increased lipid digestibility. Similarly, Gao et al. [46] reported that the addition of 5% inulin or 5% sodium carboxymethylcellulose (CMC) increased lipid digestibility in growing pigs. Agyekum et al. [47] also reported that adding 30% distillers dried grains with solubles to a growing pig diet had no effect on ileal lipid digestibility, whereas fecal lipid digestibility tended to increase. Fat or oil is typically added to fibrous diets to compensate for the energy-dilution effect of fiber. However, it has been shown that lipid digestibility increases when dietary lipid content increases [58]. Therefore, the above-mentioned studies showing that lipid digestibility increased irrespective of fiber content may be due to the increased dietary lipid content that was intended to compensate for the low energy content. Nonetheless, increasing dietary lipid content has also been reported to depress fiber digestibility [57]. Additional studies are warranted to establish the role of fiber (i.e., form and type) in lipid digestibility.
Depression in lipid digestibility has been reported to be more pronounced at the fecal level than at the end of the ileum [57]. Therefore, the values for hindgut lipid fermentation (calculated as total tract digestibility – ileal digestibility) are usually negative, indicating that dietary fiber intake may increase endogenous lipid synthesis by microflora in the hindgut of pigs. However, not all of the studies mentioned above reported a net lipid synthesis in the hindgut of pigs due to fiber ingestion. Thus, fiber ingestion did not result in net lipid synthesis in the studies by Kil et al. [58] and Gao et al. [46], whereas increased fiber intake did result in net lipid synthesis in the studies by Shi and Noblet [54], Högberg and Lindberg [45], and Agyekum et al. [47]. Kil et al. [58] and Gao et al. [46] used purified dietary fiber sources (i.e., CMC, inulin, and Solka-Floc), whereas Shi and Noblet [54], Högberg and Lindberg [45], and Agyekum et al. [47] used fiber from natural sources (i.e., cereal co-products) in their studies. Therefore, a possible reason for the discrepancy may be the fiber source used in the various studies.
《3.3. Effects of fiber on viscera organs》
3.3. Effects of fiber on viscera organs
Ingestion of high-fiber diets leads to an increased secretion of digestive juices and enzymes, as mentioned previously. This implies an increase in the workload of the secretory organs, leading to hypertrophy. In addition, in order to compensate for the low energy value of high-fiber diets, the increase in feed intake results in adaptive changes in the gastrointestinal tract [59]. For example, Jin et al. [59] observed a 33% unit increase in the rate of cell proliferation in the jejunum and colon of pigs that were fed a 10% wheat strawcontaining diet, compared with the control. These changes led to an enlargement of some or all of the organs comprising the gastrointestinal tract [60]. For example, pigs that were fed corn-soybean meal diets containing 40% alfalfa meal had heavier stomach, small intestine, large intestine, and liver weights than pigs that were fed a corn-soybean meal control diet [61,62].
Pigs that were fed diets containing barley, canola meal, and alfalfa meal showed increased per-kilogram empty body weight of the liver, cecum, and colon (on a dry matter basis) compared with pigs that were fed a casein-cornstarch-based diet [63]. In addition, pigs that were fed a diet containing 30% distillers dried grains with solubles showed increased visceral organ mass relative to the controlfed pigs [33]. However, Jin et al. [59] found that feeding growing pigs a diet containing 10% wheat straw had no effect on changes in visceral organ mass compared with the control. In the study by Jin et al. [59], pigs were fed a fibrous diet for 14 d. However, the dietary fiber level in the study by Jin et al. was lower than the fiber level in the studies mentioned above [33,61,62], which showed increases in visceral mass due to fiber ingestion in pigs. Therefore, the lack of effect of fiber ingestion on visceral mass in the study by Jin et al. [59] could be due to the inclusion level of dietary fiber and to the duration of the adaptation period employed in their study.
Studies on pigs suggest that high fiber ingestion alters intestinal epithelial morphology. For example, ingestion of a diet containing 10% wheat straw enlarged the villi and deepened the crypts in the jejunum and ileum of growing pigs compared with the control [59]. Crypt cell proliferation and depth also increased in the colon of pigs and sows that were fed diets high in fiber compared with the control [59,64,65]. Furthermore, the villus height-to-crypt depth decreased in the small intestine of pigs that were fed high-fiber diets [33,59] compared with the control, suggesting an increased crypt cell proliferation and an increased number of cells exhibiting DNA fragmentation. The effects of fiber on alteration in intestinal morphology and development depend on fiber solubility and waterholding capacity. Thus, soluble fibers are fermented to produce fermentation products—and butyrate in particular—which serve as an energy substrate for colon cell proliferation [65]. On the other hand, insoluble fibers have an abrasive effect on the intestinal wall, as they scrape the mucosa [66]. This abrasiveness may cause an increase in mucin production and in the rate of intestinal mucosal cell turnover.
《3.4 Effects of fiber on energy metabolism》
3.4 Effects of fiber on energy metabolism
The effects of fiber on the energy metabolism of pigs have been studied extensively. Results from such studies show a negative effect of fiber ingestion on the energy metabolism of pigs. Several mechanisms have been adduced to explain the negative effects of fiber on the energy metabolism of pigs. Firstly, and as mentioned previously in this review, dietary fiber may decrease the digestibility of starch, lipid, and protein, which are energy-yielding substrates, resulting in decreased energy digestibility; thus, fiber may reduce the amount of energy retained for edible pork production.
The second mechanism to explain the negative effects of fiber relates to the increased visceral organ mass that is caused by the ingestion of high-fiber diets, as mentioned previously. The visceral organs that are drained by the portal vein and liver (collectively termed the splanchnic tissues) are metabolically active tissues; thus, they require a substantial proportion of the whole-body energy (and nutrient) supply for their maintenance, although they represent less than 15% of the total body mass [67]. Indeed, Yen et al. [68] showed that the portal vein alone accounts for nearly 25% of the whole-body energy demand (measured in terms of the amount of oxygen, O2, consumed) in pigs. Furthermore, the splanchnic tissues account for nearly 45% of the whole-body energy demand in pigs [67].
Sodium-potassium adenosine triphosphatase (Na+/K+-ATPase) activities and protein turnover have been reported to be responsible for the high energetic demands of the splanchnic tissues [69,70]. A study by Kelly et al. [71] provides evidence to implicate these two processes in the energy demanded by these organs. In that study, Na+/K+-ATPase and protein synthesis activities were estimated to account for 15%–41.5% and 27%–31% of oxygen consumption by the visceral organs, respectively. Thus, it follows that changes in the portal vein mass due to high fiber ingestion would exert an additional energy demand for maintenance. For example, Yen et al. [72] reported that growing pigs that were fed a (high-fiber) diet based on barley-canola meal had an increased portal vein mass and an increased maintenance energy requirement (measured as fasting O2 consumed by the portal vein and by the whole-body animal) compared with pigs that were fed a (low-fiber) diet based on casein and cornstarch. However, Agyekum et al. [33] did not observe increased whole-body fasting O2 consumption in growing pigs that were fed a corn-soybean meal diet containing 30% distillers dried grains with solubles, in spite of an increased portal vein mass relative to pigs that were fed the control diet. The smaller sample size (four pigs per treatment) in the study by Agyekum et al. [33] may partly explain the lack of effect shown by the high-fiber diet on the maintenance requirement in the pigs. Nonetheless, the increased energetic and nutrient demand of the portal vein in pigs due to high fiber intake reduces the amount of energy and nutrients available for the growth of tissues of economic importance, such as muscle. Indeed, the ingestion of a fiber-rich diet was shown to reduce pig carcass weight in several studies, albeit with similar slaughter body weights [33,73,74].
The third mechanism by which the intake of a high-fiber diet negatively affects energy metabolism relates to the increased endogenous nitrogen and amino acid production that occurs via increased secretions of digestive juices and enzymes, which are due to fiber ingestion. This result is because the increased production of endogenous nitrogen and amino acids is associated with increased gut protein turnover, that is, increased transport of the products of digestion and blood flow through the digestive tissues [75]. However, these processes demand a substantial amount of energy, which leads to an increase in the maintenance energy requirement [3], and thereby reduces the amount of energy retained for edible pork production. In addition, increased endogenous nitrogen and amino acid production, coupled with reduced reabsorption and pre-cecal amino acid digestibility (as discussed previously), leads to an increased amino acid maintenance requirement [41,51] and thereby decreases amino acid availability for growth and production.
However, fiber can contribute to the energy supply of the pigs upon fermentation in the hindgut. In this case, the amount of energy that is contributed varies depending on feed ingredients, fiber type, and interactions with other dietary factors [10]. Upon fermentation, the volatile fatty acids produced are metabolized through a series of anaerobic energy-yielding reactions to produce adenosine triphosphate (ATP), which is used to produce energy [10]. Indeed, reports [67] suggest that the absorbed volatile fatty acids can supply up to 28% of the maintenance energy requirement of pigs. Acetate, propionate, and butyrate produce the largest quantities of energy. The numbers of moles of ATP produced by acetate, propionate, and butyrate are 10, 18, and 28, respectively, and the hydrolysis of 1 mol of ATP into adenosine diphosphate (ADP) releases 7.3 kcal (1 kcal = 4.184 kJ) of energy. Therefore, the potential energy that is absorbed upon fiber fermentation (in the case of cellulose) can be approximately 70% of the energy that is produced when glucose is oxidized. However, the amount of energy that is available to the pig for productive functions from fiber fermentation is low as a result of the energy losses due to methane and hydrogen (H2) production, as well as to the heat of fermentation [10].
《3.5. Effects of fiber on fermentation and intestinal health》
3.5. Effects of fiber on fermentation and intestinal health
Because pigs do not secrete the enzymes that are required to hydrolyze dietary fiber, the disappearance of dietary fiber in the digestive tract of pigs is ascribed to microbial fermentation. Fermentation occurs mainly in the hindgut, that is, the cecum and colon, due to the dense microbial populations in this region of the gut, although considerable fermentation can happen before the end of the ileum [3,15,76]. Furthermore, different fiber types have different susceptibility to microbial fermentation; thus, soluble fiber generally has a faster fermentation rate than insoluble fiber. Indeed, it has been shown that the fermentation of soluble fiber occurs in the proximal colon, whereas the fermentation of insoluble fiber is gradual and can be sustained until the end of the colon [77]. It has also been reported that the addition of insoluble fiber to pig diets may lead to low or insignificant microbial fermentation in the small intestine, although fermentation remains low in the hindgut [15]. Therefore, it follows that diets containing a high amount of insoluble fiber may hamper fiber fermentation in pigs. For example, Urriola and Stein [39] and Gutierrez et al. [34] reported that the addition of up to 30% distillers dried grains with solubles and 40% corn bran with solubles, respectively, to diets based on corn-soybean meal reduced the total tract and hindgut disappearance of total dietary fiber and of NDF in growing pigs compared with pigs that were fed diets based on corn-soybean meal. Nonetheless, fiber fermentation in sows is greater than in growing pigs owing to the longer digesta retention time in the hindgut and to the well-developed gut capacity in sows [37,54].
Dietary fiber fermentation by the gut microflora results in the production of volatile fatty acids—mainly acetic, propionic, and butyric acids—along with carbon dioxide (CO2), H2, and methane gases [3], which, upon absorption, contribute to the dietary energy supply [78] as previously discussed. It has also been suggested that the volatile fatty acids are beneficial to intestinal development and gut health in pigs [10,76]. For example, butyrate is used as an energy source for colonic cell proliferation. In addition, propionate and a certain amount of butyrate are used for gluconeogenesis, and nearly two-thirds of acetate is metabolized in the muscle cells [79].
As previously discussed, when fiber is fermented, the microbial fermentation products can promote the proliferation of the mucosal epithelium by increasing gut length, mass, and villus height. This in turn increases the intestinal surface area to increase the absorptive capacity. In addition, the products of fiber fermentation are used to promote bacterial growth [3,78,79]. Therefore, dietary fiber ingestion not only influences nutrient utilization, but also influences other physiological functions in the pig gut. In this context, some components of dietary fiber have been reported to confer benefits to pig gut health via the promotion of beneficial intestinal bacterial proliferation and the prevention of colonization of opportunist pathogens. Different fiber sources influence bacterial proliferation differently. For example, the fermentation of resistant starch influences the proliferation of bifidobacteria [80], whereas pectin fermentation stimulates lactobacilli proliferation [81]. The fermentation of cereal grains with high levels of insoluble non-starch polysaccharides stimulates the growth of cellulolytic Ruminococcus-like bacteria and xylanolytic Clostridium-like bacteria [82,83].
However, the results of studies on the effects of dietary fiber on the intestinal health of pigs have been contradictory. For example, some studies link soluble fiber to an increased risk of post-weaning colibacillosis and swine dysentery [84–87]. The main issue is that the soluble fiber increases digesta viscosity and undigested nutrients and delays digesta passage rate; thus, it provides nutrients and a milieu that are conducive to pathogen proliferation and colonization, which hampers animal health. Conversely, other studies [88] reported beneficial effects of soluble fiber in protecting against enteric diseases in pigs. Insoluble fiber, on the other hand, has been linked with reduced colonization of the gut by pathogenic bacteria and reduced severity of enteric infections [89–92] because insoluble fiber decreases digesta transit time and thereby decreases the time available for pathogens to proliferate. However, not all studies support the ability of insoluble fiber to promote intestinal health (Table 1) [84,85,88,89,91,93–97]. Nonetheless, it is recognized that the detrimental effect of soluble fiber on intestinal health is due to its viscosity and not to fermentability, and that combining soluble and insoluble fiber in diets produces a superior response on indices of gut health [4,93,98,99]. However, it is difficult to establish the recommended level of dietary fiber to confer health benefits on pigs, since it ultimately depends on the feed ingredients to be used for diet formulation and on the fact that a high fiber inclusion rate can hamper nutrient utilization and pig performance.
《Table 1》
Table 1 Results of studies evaluating the effects of the type of non-starch polysaccharides on performance and gut health of pigs.
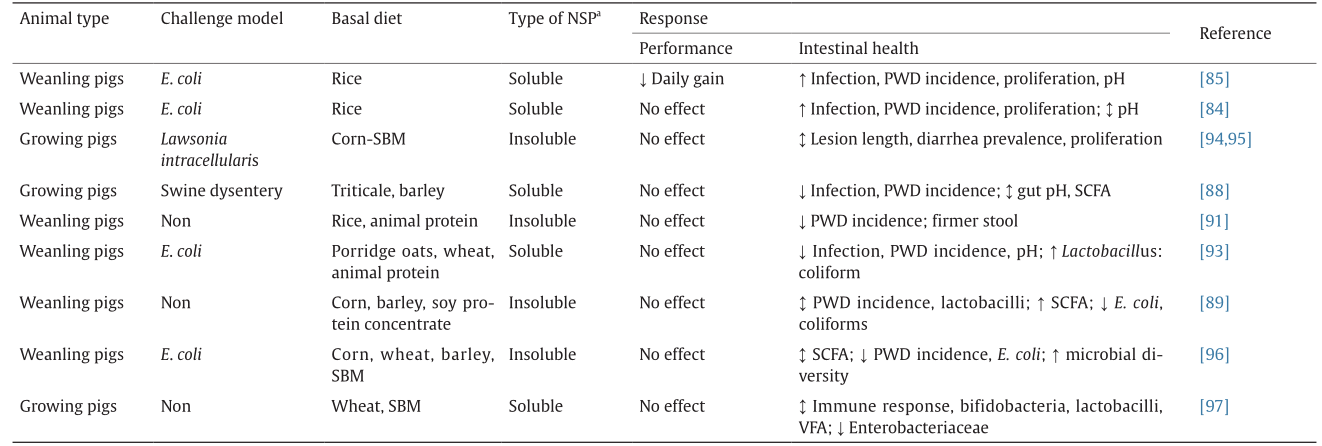
E. coli: Escherichia coli; PWD: post-weaning diarrhea; SBM: soybean meal; SCFA: short-chain fatty acids; VFA: volatile fatty acids. The symbol ↑ indicates an increase in response criteria, ↓ indicates a decrease in response criteria, and ↕ indicates no effect on response criteria.
aNSP: non-starch polysaccharides; NSP-containing diets are compared with non-antibiotic control diets for the entire experimental period; significance is defined as P < 0.05.
《3.6. Effects of fiber on gestating sow behavior》
3.6. Effects of fiber on gestating sow behavior
Restricted feeding is a common practice in gestating sows to prevent excessive body weight gain and ramifications on locomotion and reproductive functions. Although this feeding practice ensures that sufficient nutrients are provided to meet the maintenance and reproductive needs of sows, it does not provide enough feed to achieve satiety for the sow. A lack of satiety due to restricted feeding has been reported to result in aggression and stereotypies [100], which are of great welfare and production concern in individual or group-housed gestating sows. Therefore, fibrous ingredients are often incorporated into pregnant sow diets to reduce the sensation of hunger and to overcome the aggression and behavioral problems associated with restricted feeding [9]. The beneficial effects of high-fiber diets have been ascribed to their ability to delay gastric emptying [101] and increase swelling of the stomach content and fermentation products [49,102].
Fiber type or inclusion level does not influence the immediate post-feeding behavior of sows, as the feeding duration and physical activities remain the same [9]. However, the long-term effectiveness of high-fiber diets on satiety and sow behavior depends on the physicochemical properties of the dietary fiber. Soluble fiber sources are able to reduce physical activity for several hours after feeding because they can delay and sustain nutrient absorption for longer periods [9,49,102]. Soluble fiber can also increase volatile fatty acid production, which can serve as a source of energy when the glucose supply to the gut decreases. In addition, volatile fatty acids can influence the production of certain gut-secreted hormones, such as ghrelin, glucagon-like peptide 1, and peptide YY, which are associated with energy and lipid metabolism and which prolong the feeling of satiety [103]. Therefore, soluble fiber has a greater effect on satiety in gestating sows than insoluble fiber, owing to its greater ability to modify the production of gut hormones that are involved in satiety, lipid metabolism, and energy metabolism.
《4.Improving the feeding value of high-fiber diets》
4.Improving the feeding value of high-fiber diets
Feed-processing techniques such as mechanical and hydrothermal processing can be used to alter the physical properties of the diet in order to improve the solubility of feed ingredients that are rich in insoluble fiber [104,105]. These technologies can be employed along with a diet that is rich in insoluble fiber in order to examine their effects on gestating sow welfare and behavior.
Several strategies have been employed to improve the nutritive value of high-fiber diets fed to pigs, such as formulating diets based on net energy or available nutrient basis, particle size reduction, pelleting, and the use of exogenous enzymes. Such strategies have been the object of several excellent reviews. Exogenous enzyme application has been the most widely used approach to ameliorate the adverse effects of high-fiber diets on nutrient utilization in pigs. The use of this application is based on the premise that pigs do not secrete the enzymes that are required to degrade fibrous components; thus, supplementing diets that are rich in fiber with appropriate enzymes can aid in this regard. Several enzyme preparations to degrade the fiber in pig feeds have been made commercially available. Among the exogenous fiber-degrading enzymes, xylanase and β-glucanases are the most widely used in high-fiber pig diets because arabinoxylans, cellulose, and β-glucans are the most predominate non-starch polysaccharides in high-fiber pig diets [2,106]. Xylanase and β-glucanases are often used together or in combination with other commercially available carbohydrases (e.g., β-mannanase, α-galactosidase, and pectinase) as well as exogenous protease and phytase in order to effectively hydrolyze high-fiber diets and thereby increase nutrient utilization by pigs [106].
The application of exogenous enzymes has been studied extensively in swine nutrition. However, inconsistent results have been obtained regarding the effects of exogenous enzymes on nutrient utilization and growth performance in pigs that are fed high-fiber diets (Table 2) [107–113]. Factors such as variation in dietary ingredient composition, variation in experimental conditions, and enzyme characteristics (i.e., source, dose, and matrix) have been put forward to explain inconsistent study results regarding the effects of enzymes in pig trials [2,114].
《Table 2》
Table 2
Effects of supplementing fiber-rich diets with carbohydrase complex on nutrient digestibility and performance in pigs.
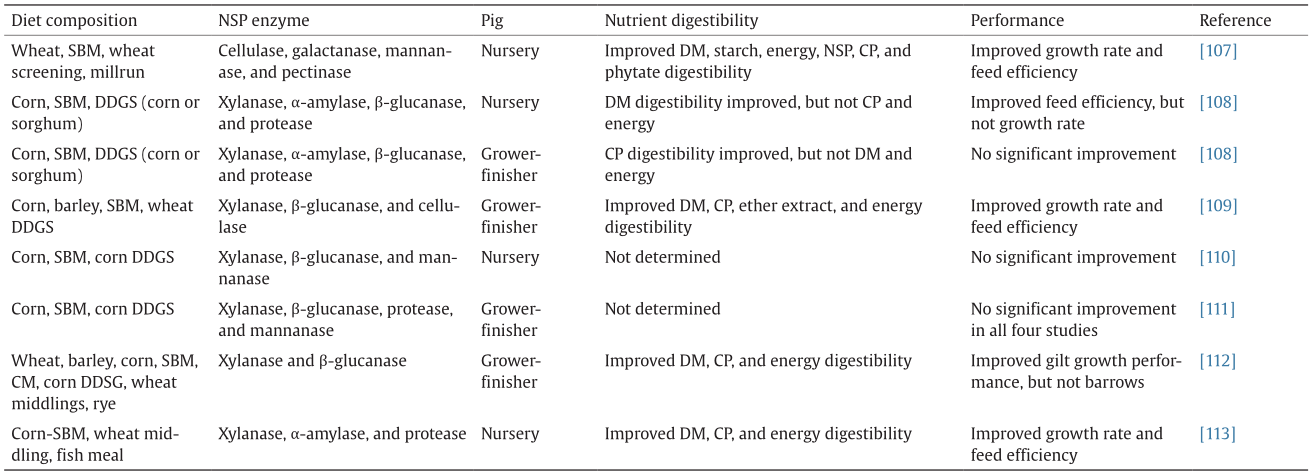
CM: canola meal; DDGS: distillers dried grains with solubles; CP: crude protein; DM: dry matter.
The mechanism by which supplemental enzymes influence the metabolic response and thus the digestive physiology of pigs that are fed high-fiber diets is still not clearly understood. For example, growth is commonly used to assess the efficacy of enzymes in pigs that are fed high-fiber diets; however, growth relies on the interplay of biological and biochemical processes. Thus, specific processes within an animal must be suppressed or enhanced in order for an animal to grow. Therefore, it is necessary to elucidate how exogenous enzymes influence these processes to elicit growth in pigs that are fed high-fiber diets. Furthermore, studies on the effects of enzymes on high-fiber diets have concentrated on activities within the gut lumen (e.g., nutrient digestibility and digesta pH and viscosity) without considering the consequences of feed enzymes on nutrient absorption patterns and the growth-related endocrine system; more specifically, these studies do not consider the effect of feed enzymes on the secretions of important gut hormones that are involved in increasing pig performance. However, the use of surgically modified pig models (e.g., portal vein catheterization) and slaughter techniques permits digestion to be studied in different segments of the digestive system, along with the associated metabolic and physiological effects. The data generated from such studies can assist in explaining the lack of improvement in pig growth even in the presence of improved nutrient digestibility when enzymes are added to high-fiber diets, and can thus assist in the design of strategies for effective nutrient utilization in pigs that are fed high-fiber diets.
《5.Conclusion and perspectives》
5.Conclusion and perspectives
Fibrous co-products present an opportunity to reduce the cost of raising pigs and thus enhance profitability. However, incorporating such co-products into pig diets will inevitably increase the dietary fiber content. Ingestion of diets that are rich in fiber has been associated with impaired nutrient utilization leading to growth depression. On the other hand, dietary fiber has received considerable recent attention owing to its ability to influence pig gut physiology and gut health, and sow welfare. However, reports in the literature regarding the effects of fiber on pig growth responses and gut health have been contradictory. These contradictions are due to differences in fiber sources, type, and inclusion level, and in the analytical methods used to measure dietary fiber in the literature. Additional research is required to clarify the effects of fiber source, type, and inclusion level on pig performance and gut health.
Several approaches can be used to improve the nutritive value of high-fiber diets in pigs. Fiber-degrading enzymes are the most researched and widely used approach to offset the negative effects that are associated with the ingestion of fiber-rich diets, and allow a wide range of fibrous feed ingredients to be used in swine feeds. However, the effects of fiber-degrading enzymes in fiber-rich pig feeds have not been consistent among studies; therefore, additional studies are warranted in order to thoroughly investigate the inconsistencies. One way of addressing these inconsistencies would be to investigate the influence of fiber-degrading enzymes on the expression of genes that are associated with intestinal integrity and nutrient transport, because such genes relate to the normal function of the intestinal epithelial cells and to the ability of the intestine to absorb nutrients from the gut lumen. Lastly, feed-processing techniques can be employed to improve the solubility of feed ingredients that contain high levels of insoluble fiber. Such technology could be applied to increase the use of insoluble fiber in diets for gestating sows in order to improve sow welfare and behavior.
《Compliance with ethics guidelines》
Compliance with ethics guidelines
Atta K. Agyekum and C. Martin Nyachoti declare that they have no conflict of interest or financial conflicts to disclose.











 京公网安备 11010502051620号
京公网安备 11010502051620号




