

《1. Introduction》
1. Introduction
Efficient biological nitrogen removal has attracted much attention in recent years [1–3]. The wondrous discovery of bacteria performing complete oxidation of ammonia (so-called comammox, CMX) to nitrate (NO3– ) via nitrite (NO2– ) has challenged the understanding of nitrogen removal. CMX bacteria have been widely reported in both natural [4–7] and engineered systems [8–11], implying their ubiquity as well as their contributions in various nitrifying habitats. CMX bacteria, having a high growth yield propensity [12], are well fitted to water treatment plants (WTPs) and wastewater treatment plants (WWTPs), particularly plants that have low oxygen accessibility and low cell washout rates (such as biofilms). Thus, revalidation of nitrifying environments has attracted much attention, which likely provides meaningful insights into the understanding of CMX bacteria, particularly clade A Nitrospira sp. [13–17]. While producing a high-quality nitrate solution (to be used as hydroponic fertilizer) from organic nitrogen using a dual reactor system, Sato et al. [18] found a high abundance of CMX Nitrospira-related operational taxonomic units (OTUs) in the nitrification reactor, where the expression of CMX-ammonia monooxygenase (AMO) was found to be 500 times higher than that in the canonical reactor. In a long-term study, four antibiotics (i.e., ampicillin, kanamycin, lincomycin, and trimethoprim) could successfully enrich phylogenetically distinct CMX Nitrospira in the activated sludge system [19]. Likewise, in very recent investigations, CMX bacteria were found to offer undisputable advantages in WWTPs, such as biotransformation of antibiotic [20,21] and heavy metal tolerance [22], avoiding the generation of N2O in the intermittently aerated lab-scale sequencing batch reactor [23], survivability at low temperature [24], dominance of tertiary rotating biological contactors (RBCs) biofilm samples [10], occurrence of CMX Nitrospira in urine-fed membrane bioreactors [25], and so forth, which strongly implies that CMX bacteria are fully functional in WWTP facilities.
The existence of CMX bacteria was hypothesized in 2006 [26]. The occurrence of CMX bacteria was confirmed in 2015 [7,27], although full-fledged operations of WTPs or WWTPs with CMX bacteria have not been reported. CMX bacteria have been studied from different angles, such as niche preference and differentiation [28–30], co-occurrence [31,32], cooperative and competitive interactions [11,33,34], metabolic versatility [35], biotransformation of pollutants [36], enzymology [37–39], biochemistry and physiology [40,41], and molecular detection at the clade level [42]. In addition, the response of CMX bacteria to seasonal changes and external perturbations has also been largely explored [21,43–47]. These attempts shed light on the potential vision that it is not far from now when CMX-based processes are applied in WTPs or WWTPs.
CMX bacteria have considerable disparity in many aspects, despite their uniqueness in their reaction mechanisms [38] and enzymes [27] responsible for NH3 oxidation to NO3– . For instance, not all types of CMX Nitrospira are cultivatable in vitro, which has been disclosed by emphasizing Nitrospira sp. at the subcladelevel [48]. Additionally, there is a significant difference in the responsiveness of clade A and clade B CMX to external perturbations [49]. Thus, CMX Nitrospira have unique niche requirements[13,16,30], and knowledge of these requirements helps us not only understand how CMX bacteria withstand competition from canonical nitrifiers (e.g., ammonia-oxidizing archaea (AOA), ammoniaoxidizing bacteria (AOB), and nitrite-oxidizing bacteria (NOB)) but also reveal their coexistence with canonical nitrifiers [8,32,50,51]. Importantly, the niche of CMX bacteria may raise doubt as to whether all clades and subclades of CMX bacteria have a similar type of coresident bacteria because such multispecies assemblages are the key drivers of community functions. Therefore, there should be close linkages between CMX bacterial functionalities and niche determinants. On the other hand, physiological, biochemical, and molecular studies have revealed that CMX bacteria have wide metabolic potential, such as the enhanced removal of micropollutants from wastewater by metabolic [36,52,53] and co-metabolic routes [54–58]. We believe that the above insights will aid in the deep understanding of CMX bacteria in bringing closure to engineered settings for sustainable water or wastewater treatment in nitrifying environments.
To date, the occurrence of CMX in WTPs or WWTPs has been reported widely [8,10,15,17,59–67]. Note that previous reviews have advanced the understanding of CMX in the following aspects: genomic insights [8]; nitrous oxide (N2O) emissions [64]; and enzymology and metabolic models [65]. Nonetheless, several key aspects of CMX bacteria are not yet fully understood, such as their distribution in natural and engineered systems, niche differentiation, co-occurrence, metabolic versatilities, interactions of CMX bacteria with other bacteria, reactor adaptability, stress response, and co-metabolic biotransformation. Thus, the present paper aimed to critically review these factors, and these insights would be useful in bringing CMX bacteria much closer to WTPs and WWTPs.
《2. Ubiquity of CMX bacteria》
2. Ubiquity of CMX bacteria
《2.1. Occurrence in natural and engineered systems》
2.1. Occurrence in natural and engineered systems
The distribution and abundance of CMX bacteria in different natural and engineered systems are summarized to reveal the ecology of microbial nitrification in the nitrogen cycle (Fig. 1 [48,68]). Based on the gene-encoding subunit A of the ammonia monooxygenase (amoA) phylogeny, there are two novel CMX Nitrospira representing two divergent sister clades, namely, clade A and clade B [7]. Along these lines, CMX bacteria of clade A and clade B have been detected in various ecosystems [68], implying that they are environmentally widespread. Recently, cultivatable and uncultivatable Nitrospira were found to be present in subclades A1 and A2, respectively [48]. CMX Nitrospira has been detected in different soil and freshwater ecosystems [69], WWTPs and WTPs [31], sand filters of groundwater [9], aquaculture ponds [70], and nitrifying biofilms [64], suggesting that these niches commonly serve as nitrification platforms. Although there have been limited studies, CMX Nitrospira seems to be more abundant in WTPs [9] than in WWTPs [60] and soils [71]. Additionally, within the same microenvironment, the abundance of clades A and B is normally different. For instance, clade B is more abundant than clade A in forest soils. In comparison, the abundance of clade B was 14 times lower than that of clade A in paddy soil [49]. It is noteworthy that CMX bacteria in clade B are more sensitive than those in clade A to the external environment. For example, NH4+ amendment did not show a significant impact on the abundance of clade A, while clade B abundance significantly (P < 0.05) increased with the same amendment [49]. Similarly, the presence of residual chloramine ((1.74 ± 0.21) mg·L–1) in drinking water distribution systems favored a shift toward Nitrospira-like OTUs [72]. It is worth noting that there are several primers for the identification of clades of CMX bacteria with different matching percentages (Fig. 2 [48]). For example, primers A378f show high matching (80%–90% with the amoA gene) with CMX clade A (subclade A1 and subclade A2) but low matching (< 20%) with clade B and AOB (Fig. 2), which clearly suggests that there are massive differences in the genetic potential between clade A and clade B.
《Fig. 1》
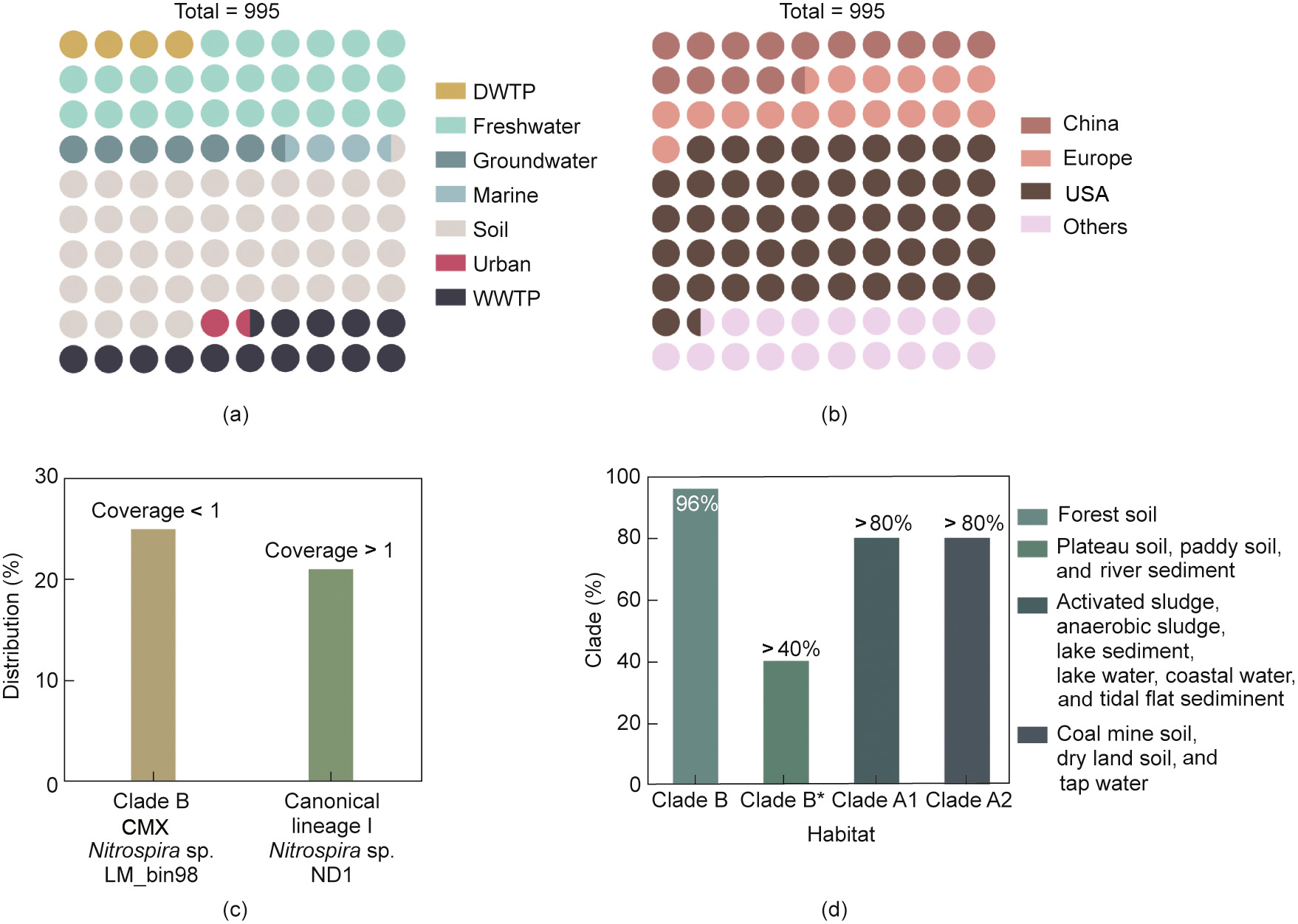
Fig. 1. Ubiquitous distribution of CMX bacteria. Distribution of Nitrospira species (a total of 68) across 995 metagenomes across (a) seven broadly defined habitats and (b) the globe, and (c) CMX Nitrospira was the most widely distributed (low abundance with a coverage < 1) than canonical Nitrospira (high abundance with a coverage of > 1) [68]. (d) Clade-level distribution of CMX bacteria in different habitats analyzed by partial nested PCR with CMX-specific primer sets [48]. DWTP: drinking water treatment plant.
《Fig. 2》

Fig. 2. Primers and their matching (%) with the amoA gene sequences of different CMX clades and AOB [48]. Sequences are extracted from metagenomic or metatranscriptomic data sets in public databases as described in the reference.
In full-scale WWTPs, CMX bacteria have been found to be an important NH3-oxidizing group [7]. A primer pair specifically designed to target the CMX amoA gene revealed that the Nitrospira nitrosa cluster accounted for 94.34% of bacteria in WWTPs [60], which was about 183-fold (as determined by quantitative polymerase chain reaction (qPCR)) more abundant than the AOB amoA gene. The dominance of CMX Nitrospira over AOB was also found in a nitrification reactor fed real municipal wastewater, where 94% of the NH3-oxidizing community at 400 d was the former group [73]. Similarly, CMX Nitrospira showed dominance over AOB Nitrospira in the organic carbon-rich nitrification–denitrification system[74]. These insights clearly suggest that CMX bacteria have solid competitive advantages over other nitrifying communities in WWTPs and particularly in WTPs, which is promising for the engineering application of mainstream CMX (Table 1 [8,12,14,41,43,63,70,75–77]). Notably, although CMX bacteria have a full suite of enzymes for both NH3 and NO2– oxidation, proper markers should be selected to characterize the presence and abundance of such enzymes. For instance, nitrite oxidoreductase (Nxr) of CMX Nitrospira is very similar to that of strictly NO2-oxidizing Nitrospira [37], which is not a suitable marker of CMX Nitrospira.
《Table 1》
Table 1 Comparison between CMX bacteria and canonical nitrifiers.

Km is described as the concentration of the substrate to which the velocity of the reaction is half maximal.
《2.2. Niche differentiation》
2.2. Niche differentiation
In each ecological niche, CMX Nitrospira shows a complex response to the distribution of resources. It has been indicated that dissolved oxygen (DO) fluctuations in water may not affect the CMX community [28], likely because CMX Nitrospira contains genes that allow these bacteria to grow even at low levels of oxygen, as determined in a comparative genomic study [29]. However, the genome of CMX Nitrospira does permit assimilatory nitrate reduction by using an external nitrite nitrogen source, which is contrary to canonical Nitrospira. Other potential niche determinants in CMX Nitrospira include levels of nitrite (activated sludge) [13], elevated N deposition (forest soils) [30], and pH-dependent NH3 availability (agricultural/forest soils) [16,30]. Nonetheless, subclades of CMX Nitrospira can achieve rapid niche divergence from other canonical nitrifiers by horizontal transfer of genes encoding NH3 oxidation [29] and hydroxylamine oxidoreductase [68]. These studies have indicated that the genomic traits of CMX Nitrospira are impacted by biotic factors [32]. Niche differentiation is an important determinant shaping microbial community structure [78]. However, there is a scarcity of information on the niche specialization, preference and differentiation of CMX Nitrospira with respect to their habitats, such as trophic states and oxygen levels [28,40]. Niche separation between CMX Nitrospira and canonical nitrifiers should be further studied by focusing on the metabolic versatilities and comparative genomics of the former community. Moreover, the niche divergence and separation of CMX Nitrospira have been explored in natural settings (aquatic [28] and soil [16,30,40] settings) and in engineered settings [73,79]. However, additional reactor-based studies are largely needed for a deeper understanding of niche differentiation of CMX Nitrospira in various facilities, including habitat heterogeneity, environmental conditions, and biointeractions.
《2.3. Functionalities at the gene level》
2.3. Functionalities at the gene level
CMX bacteria are known to coexist with different canonical bacteria, such as NOB [32], AOA [8,50], AOB [8,51], and anammox bacteria [34], which clearly implies the considerable microbial interactions of CMX Nitrospira with other canonical communities. There is a significant positive relationship between the amoA-Niño gene of CMX bacteria and selective genes of canonical nitrifiers and other bacteria (Fig. 3(a) [43]). The presence of a large number of nitrogen-cycling genes in Nitrospira inopinata (CMX) could help Nitrospira moscoviensis (NOB) utilize different metabolic functions in a freshwater mussel habitat [32]. This is because Nitrospira inopinata has more N-cycling genes than Nitrospira moscoviensis, and the subsequent metabolic sharing between the two strains is the key factor for the survival of two bacteria in the mussel habitat. Furthermore, CMX Nitrospira is known to possess a complete set of genes (urtABCDE) for a high affinity of the urea transporter system, whereas Nitrospira moscoviensis lacks a full array of these genes (having only UrtA) [32]. In addition, CMX Nitrospira has a tendency to oxidize NH3 more than NO2– under very low oxygen concentrations, which favors the cooccurrence of CMX Nitrospira and anammox bacteria [27]. In addition, the concentrations of nitrate and total nitrogen show a positive correlation with CMX clade A1 and clade A2, respectively [51]. We can expect that these clades may have different responsive behaviors to coresident canonical nitrifiers. Therefore, the cooccurrence of CMX Nitrospira should be evaluated at the subclade level to unravel the uncertainty in the cooperation and competence of CMX Nitrospira with others in nitrogen conversion systems during water and wastewater treatment [80]. It would aid in process design and operation in both WWTPs and WTPs.
《Fig. 3》
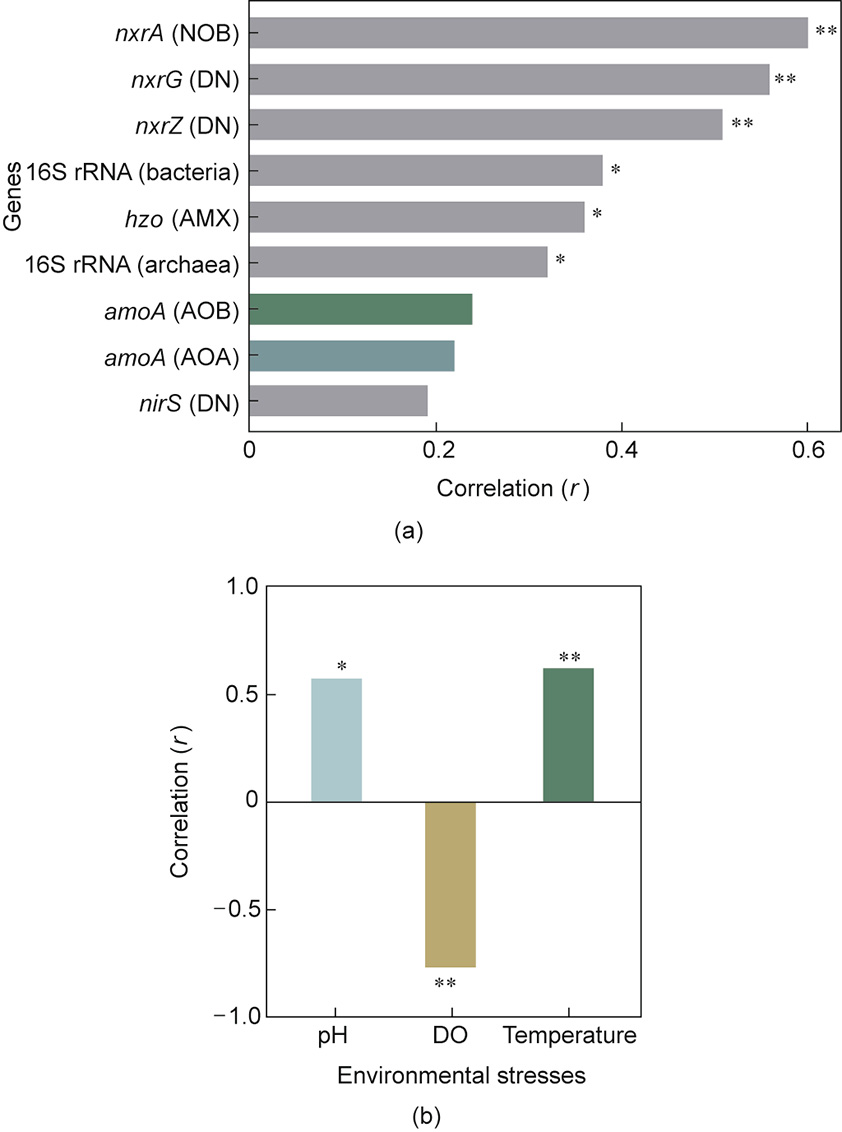
Fig. 3. Pearson relationship of amoA-Niño [43]. (a) Co-existence of CMX bacteria with other bacteria including canonical nitrifiers. Correlation between amoA-Niño and other gene appellation. (b) Correlation between amoA-Niño gene and environmental factors. AMX: anammox; DN: denitrifying bacteria; DO: dissolved oxygen; significance Two-tailed * ≤ 0.05 and ** ≤ 0.01.
In summary, CMX Nitrospira can be readily detected in natural waters, drinking water systems and oligotrophic wastewater. This is very similar to the AOA. This is likely due to the low growth rates of canonical nitrifiers in oligotrophic conditions. The high activity of CMX bacteria at low oxygen levels implies that it is possible to develop a low-cost nitrogen removal process. We should keep in mind that the boundary conditions of CMX bacteria and canonical nitrifiers in engineered systems should be determined with welldesigned experiments.
《3. Physiology and biochemical potentials of CMX bacteria》
3. Physiology and biochemical potentials of CMX bacteria
《3.1. Metabolic versatilities》
3.1. Metabolic versatilities
Some strains of CMX Nitrospira can respire on O2 or NO3– as terminal electron acceptors and oxidize different substrates, such as NH3, H2, formate, and NO2– [35]; thus, their versatility in metabolism allows these bacteria to colonize a broad range of habitats. They can oxidize NH3 by ammonia monooxygenase (AMO) and can fix CO2 by using the reductive citric acid cycle. Due to the high affinity for NH3, as confirmed by whole-cell (Nitrospira inopinata) kinetics [38], the population of CMX Nitrospira inopinata is usually higher than that of many other NH3-oxidizing microbes under oligotrophic conditions. The high NH3 affinity indicated by the halfsaturation constant (Km(app)) of a pure cultured CMX bacterium (e.g., Nitrospira inopinata) is 63 nmol·L–1 , which is 4- to 2500-fold below the values reported for AOB [41]. Even a much lower Km value (i.e., 49 nmol·L–1 ) for NH3 was observed for clade A CMX bacteria [41]. Nonetheless, CMX bacteria show a lower affinity toward NO2– than other NOB [41]. Similar to NH3, cyanate can also be utilized by CMX Nitrospira during complete nitrification [74]. Furthermore, CMX Nitrospira is likely to adapt to oligotrophic conditions [35], which assists CMX bacteria in outcompeting AOA in oligotrophic environments [40]. It is also worth noting that if there is any overlap in Km(app) values, competition exists between CMX Nitrospira inopinata and other canonical nitrifiers (e.g., AOA) in oligotrophic and low-NH3 available habitats, although there is a higher abundance of the amoA gene in CMX Nitrospira than in AOA or AOB amoA. Surprisingly, changes in soil pH ranging from 4.0 to 9.0 had a mild impact on the abundance of amoA gene copy numbers of both clades A and B of CMX bacteria, although the abundance of clade A was higher than that of clade B at each measured pH value [40]. Nevertheless, pH, DO, and temperature have a significant influence on CMX bacteria, as revealed by amoA-Niño gene abundance (Fig. 3(b)). CMX Nitrospira is able to thrive successfully in WWTPs with low and fluctuating urea concentrations[40], attributed to the presence of genes encoding urease proteins, high-affinity urea transporters (urtABCDE), urea carboxylaserelated transporters (uctT), and outer-membrane porins (fmdC), as revealed by metagenomic analysis [7,27]. Recently, it was found that the release of N2O by CMX microbes during nitrification is comparatively lower than that by AOB [6] because of the lack of NO reductases in the genome of Nitrospira inopinata. In fact, nitrifying biofilms of canonical nitrifiers have higher N2O emissions[64]. Therefore, less emission of N2O by CMX helps to decrease the cost associated with improved plant design and operation to mitigate N2O emissions during wastewater treatment. In addition, the metabolic potentials of CMX Nitrospira due to the presence of [NiFe]-hydrogenases (group 3b) can allow these bacteria to exhibit distinct roles [12], such as ① production of H2 by reoxidizing NAD(P)H and concurrent maintenance of cellular redox balance during fermentation, ② oxidation of H2 to provide electrons for CO2 reduction, and ③ reduction of elemental sulfur or polysulfide to H2S. Furthermore, it is not clear how CMX bacteria switch between NH3 and NO2– oxidation modes. When Nitrospira phylotypes were fed inlabeled NH4+ and NO2– columns (composed of rapid gravity sand filter material), CMX Nitrospira did not show a preference for only oxidizing external NO2– [81], which was in line with Nitrospira inopinata [7,27]. It is worth noting that ClO3– and NO-chelating compounds (e.g., 2-phenyl-4,4,5,5-tetramethylimida zoline-3-oxide-1-oxyl (PTIO)) exert inhibitory effects on NH3 oxidation by CMX Nitrospira [81] and Nitrospira inopinata [6], respectively. In fact, reduction of ClO3– generates toxic ClO2– , which is toxic to CMX bacterial metabolism (including NH3 oxidation). The metabolic potentials of different clades of CMX bacteria were compared with AOB and AOA, and their relevance to WWTPs is presented in Table 2 [8,82–91]. It is also worth noting that there is possible lateral gene transfer in CMX Nitrospira, by which these bacteria obtain certain additional metabolic functions [10,14,92].
《Table 2》
Table 2 Metabolic potential of CMX bacteria and their canonical nitrifiers.

+++ means function is encoded in all genomes; + means function is encoded in some to most genomes; means function is not encoded.
《3.2. Enzymology》
3.2. Enzymology
Eight-electron oxidation reactions are mediated by CMX bacteria during the complete oxidation of NH3 to NO3– [38]. CMX-mediated nitrification is potentially dependent on the expression of genes, such as amo, hao, and nxr [27]. A study based on metagenome-assembled genomes confirmed the potential of CMX in hydrolyzing urea to NH3 by using the ure genes in the genome of CMX Nitrospira with RBCs [10]. These genes have also been reported in urea-enriched cultures of clade A CMX Nitrospira [29]. In contrast, clade A CMX Nitrospira cannot utilize formate as an alternative electron donor since fdh genes are absent in these bacteria, whereas clade B CMX Nitrospira are known to possess fdh genes [10]. In addition, some CMX Nitrospira can potentially use dihydrogen/protons as alternative electron donors/acceptors, which is attributed to genes such as hyb and hyd encodinssg group 3b [Ni–Fe] sulfur-reducing hydrogenase [11]. By lateral gene transfer, CMX Nitrospira acquire not only cyanase activity [10] but also two toxin–antitoxin genes adjacent to cyanase genes [14]. Nonetheless, horizontal gene transfer (also called vertical inheritance) helps in the recruitment of diverse genes from nonammonia oxidizers to facilitate the overall ammonia oxidation pathway [92]. In fact, CMX bacteria inherently have cyanase activity [17]; therefore, these bacteria utilize diverse organic nitrogen compounds in addition to free ammonia as the substrate. It is noteworthy that the other metabolic versatilities of CMX Nitrospira bring these bacteria much closer to WWTPs. Han et al. [36] found that CMX Nitrospira inopinata was able to biotransform a wide range of micropollutants, where carbendazim was exclusively biotransformed by CMX Nitrospira inopinata, where there was a supply of ammonia rather than nitrite as the energy source. These results strongly imply that the AMO of CMX Nitrospira inopinata has a much higher affinity toward ammonia than that of AOA- or AOB-Nitrospira. Additionally, the enzymatic system of CMX Nitrospira inopinata enables these strains to emit lower levels of NOy (nitrous acid (HONO) + nitric oxide, and NO + nitrogen dioxide (NO2)) [93], which is a critical emission in global warming and environmental chemistry. In short, the presence of these functional enzymes can endow CMX bacteria with various metabolic potentials.
《3.3. Interactions of CMX bacteria with other bacteria》
3.3. Interactions of CMX bacteria with other bacteria
Interactions between CMX bacteria and other bacteria (e.g., canonical nitrifiers) would greatly impact the design and optimization of WWTP facilities. Nitrospira inopinata showed significant positive correlations with NOB rather than AOA and AOB [43], implying that CMX-mediated NO2– oxidation is more active than NH3 oxidation. Similar results of CMX bacteria and NOB interactions have also been found in recirculating aquaculture systems [31]. Although these positive correlations are often present in other combinations (Nitrospira inopinata-like CMX and denitrifying microbes [32]), we are far from fully understanding the multiple correlations among CMX bacteria and canonical nitrifiers due to complex environmental factors. In particular, in natural circumstances, it is difficult to tackle biological factors in engineered systems. Cooperation among partial nitrifiers, CMX and anammox bacteria achieved the removal of 98.82% of NH4+ -N in a sequencing batch reactor [34], suggesting that CMX bacteria can play cooperative roles in concurrent high nitrogen removal. In particular, under low oxygen conditions, CMX bacteria oxidize NH3 to NO2– , which favors interactions between CMX and anammox [11]. Cooperation between AOB and CMX Nitrospira has been found to ensure efficient Cu-stimulated nitrification in groundwater-fed rapid sand filters [45]. To understand the community behavior of CMX bacteria, research toward quorum sensing (QS) in CMX has already been undertaken, for example, CMX bacteria contain genes responsible for the synthesis of acyl homoserine lactones (AHLs) and extracellular polymeric substances (EPS) [77]. In fact, QS has been identified in pure [94] and mixed cultures [95] of canonical nitrifiers, as well as in autotrophic nitrification/denitrification biofilms [96]. The biofilm formation potential of CMX bacteria is well regulated by the genes present in ABC transporters (ko02010) and two components (ko02020) [97], corroborating that CMX can prefer to grow in biofilms. In fact, the CMX process has already been identified in biofilm reactors [98], but the molecular details of their QS in these settings are not yet known [99].
Overall, metabolic versatilities are the unique feature of CMX Nitrospira because of the expression of various functional genes in CMX cells, implying that CMX Nitrospira can grow both in sewage and industrial wastewater. In addition, the low generation of N2O by CMX Nitrospira over canonical nitrifiers suggests that the application of CMX would aid in developing sustainable nitrogen processes in the near future.
《4. Behavior of CMX bacteria in engineered systems》
4. Behavior of CMX bacteria in engineered systems
《4.1. Reactor adaptability》
4.1. Reactor adaptability
CMX Nitrospira possesses distinct physiological characteristics, such as a lower Km value for NH3, a higher μmax, a higher mg protein per mole NH3, and a higher O2 consumption rate in terms of mole O2:NH3 than other canonical nitrifiers [8]. These features enable CMX Nitrospira to have great resilience in WTPs and WWTPs, where fluctuations of water/wastewater compositions and ambient conditions often occur. A combination of molecular approaches (i.e., qPCR, 16S rRNA gene sequencing and metagenomics) confirmed the dominance of CMX Nitrospira over AOA and AOB in the tertiary rotating biological contractors of a municipal WWTP [10], indicating that CMX bacteria are ubiquitous in tertiary WWTP systems. Additionally, the presence of CMX bacteria in the bioreactor could provide a way to understand the impacts of various environmental factors on the distinct spatial and temporal distributions of CMX [59]. Nevertheless, there is a significant contribution of CMX to nitrification [100]. The molecular analysis of samples collected from different engineered systems revealed that the ratio (%) of CMX-coding DNA sequences (CDS) and total CDS were between about 0.3% and > 0.6% (Fig. 4(a) [15]). The dominance of CMX Nitrospira sp. was also found in a dechlorinated reactor (Fig. 4(b) [72]), where the sequences comprised 7.2% to 27.4%, and these bacteria were the major candidates responsible for nitrification in the dechlorinated reactors. These studies clearly indicate the adaptability of CMX bacteria to reactor systems.
《Fig. 4》

Fig. 4. Reactor adaptability of CMX bacteria. (a) Ratio of CMX coding DNA sequences (% CMX CDS) and total CDS in full-scale WWTPs surveyed in different countries [15]. Type of feed (1SS, 2MS) and biomass (agranule, bbiofilm, c mixed liquor, doverflow, eunderflow, fALT (mixed liquor), ginoculum, and hActivated sludge). (b) Dominance (mean with standard deviation) of Nitrospira sp. in a dechlorinated reactor, showed in a Scatter plot with bars [72]. MBBR: moving bed biofilm reactor; BNR: biological nutrient removal.
It is important to note that NH3 concentrations alone are not a suitable determinant for the abundance of CMX Nitrospira in engineered systems. Despite high ammonium concentrations, low [101] and high abundances [61] of CMX bacteria have also been reported in sludge samples. It is difficult to assess the abundance of CMX bacteria in reactors unless the studies examined substrate ranges, oxygen requirements, growth rates and yields, and biofilm formation propensities. Factors affecting the presence and dominance of clade A and clade B are not yet well unknown [10]. Roots et al. [73] found higher ammonium removal rates (about 59 mg-N·(L·d)–1) by CMX Nitrospira at a DO of 0.2–1.0 mg·L–1 in a mainstream nitrification reactor fed real municipal wastewater (operated for > 400 d) compared with a nitrifying conventional activated sludge reactor with DO = 3–5 mg·L–1. This clearly suggests that CMX-driven low-DO nitrification is an alternative to conventional high-DO-driven nitrification. Furthermore, in a recent investigation, it was found that CMX bacteria were well adapted in a reactor fed either NH3 or NO2– as a sole nitrogen source [102]. However, the nitrification rates of CMX bacteria were nearly 2-fold higher when dosed with NH3 than when dosed with NO2. This finding can largely strengthen the application of CMX-based processes. Nonetheless, the ratio of either CMX bacteria to canonical nitrifiers, or clade A to clade B, was significantly affected by nitrogen–phosphorous–potassium (N–P–K) fertilization (Fig. 5 [103]). A recent study also revealed that, in comparison with other ammonia-oxidizing microbes, CMX bacteria were more severely inhibited by free ammonia in a partial-anammox bioreactor [104].
《Fig. 5》
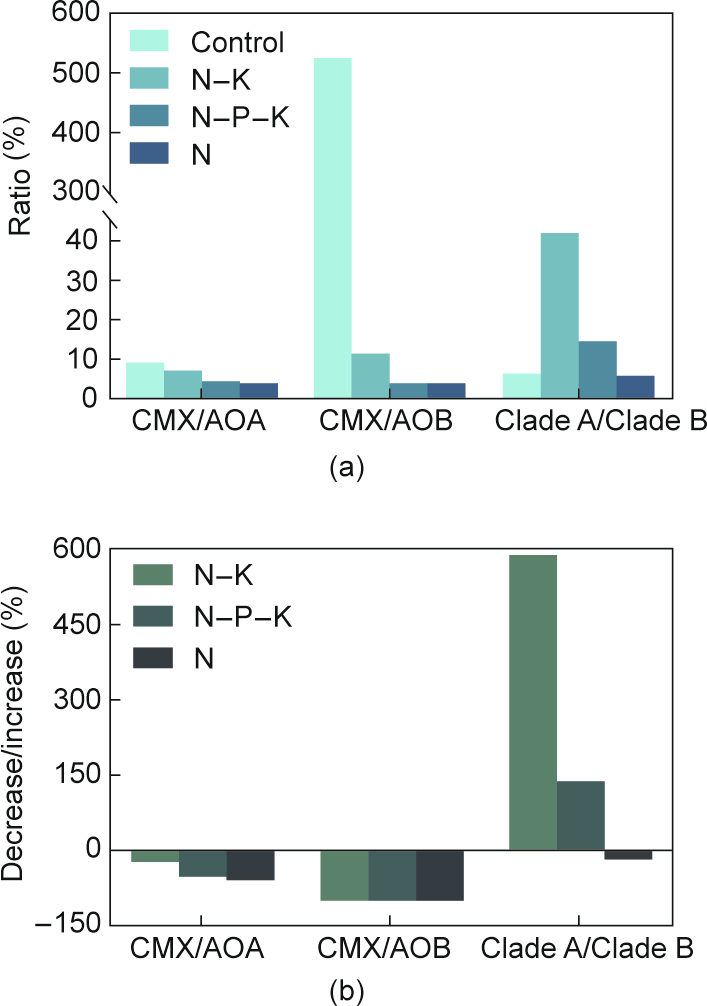
Fig. 5. Influence of nutrient amendments on the CMX bacteria and canonical nitrifiers [103]. (a) Ratios of amoA gene copy numbers under different long-term fertilization regimes. (b) Per cent decline or increment in the ratios of amoA gene copy number over untreated control.
Furthermore, long solid retention times (SRT > 10 d) and attached growth mode are favorable conditions for the growth of clade A CMX bacteria [11], and SRT was found to be the key determinant for the prevalence of CMX bacteria. Certainly, a long SRT has a significant impact on the microbial community in the reactor system, for example, biofilm thickness [105], microbial biodiversity [106], biomass concentration [107], and microbial volatile acid production [108]. Moreover, it was found that Nitrospira suppressed NO2– accumulation and inhibited N2O generation under a long SRT [109]. It is worth noting that enrichment of CMX Nitrospira is feasible in nitrifying granules, and these enriched cultures showed satisfactory efficiency in nitrogen removal [110]. Interestingly, mucopolysaccharides from invertebrates can affect CMX bacteria in a natural setting (e.g., coastal sediments), that is, they can stimulate NO3-oxidizing functional groups [111]. Recently, it was also found that CMX Nitrospira exhibited efficient nitrogen removal in engineered systems at low DO and NH4+ and high SRT [73]. These insights provide potential clues for the optimization of bioreactor configuration and operating parameters. Furthermore, quantitative PCR and PCR cloning experiments revealed that CMX bacteria can better adapt to biofilms than to suspended sludge in WWTPs [62]. Furthermore, according to phylogenetic analysis of amoA genes, it was found that uncultured CMX Nitrospira are predominant in biofilms [62]. As current WWTPs are mainly based on biofilm and activated sludge processes, these findings are of great importance for the operation and design of WWTPs. This may be a potential reason why CMX are ubiquitous in WTPs or drinking water distribution systems where bacteria can normally be present in the form of biofilms.
《4.2. Stress response》
4.2. Stress response
CMX Nitrospira can firmly withstand physically and chemically induced stresses. For instance, Nitrospira was constantly detected in activated sludge when subjected to NH3 starvation stress, where the DNA and complementary DNA (cDNA) of Nitrospira were not affected by fluctuations in temperature throughout the starvation experiments [44]. Some gene-encoding enzymes (e.g., superoxide dismutase, catalase, and several peroxidases) can protect bacteria against reactive oxygen species (ROS) [17]. CMX bacteria can adapt well to a common disinfectant, for example, peracetic acid (PAA), in aquaculture systems. PAA (application rate 1.1 mg·L–1 twice per day) showed adverse effects on the gene copy numbers of CMX bacteria after exposure for eight weeks, but these effects gradually decreased at week 13 [112]. This strongly indicates the potential adaptability of CMX bacteria to long-term chemically induced stress. Such an adaptive response of CMX bacteria was also found when exposed to long-term fertilization [46]. There was an alleviation of tetracycline (TC) stress on the nitrifying population in the presence of a low Cu(II) content. The abundance of functional genes related to nitrification and denitrification was adversely affected at 20 mg·L–1 TC, but these effects were diminished in the presence of a mixed stress of TC (20 mg·L–1) and Cu(II) (10 mg·L–1) [113] due to the decreased TC inhibition on enzyme activity at low Cu(II) content.
Importantly, the adaptability of CMX bacteria has been observed to switch between lifestyles in fluctuating environments. For example, CMX bacteria trigger the biosynthesis and degradation of intracellular compounds, such as polyphosphate, glycogen and polyhydroxyalkanoates, as a part of their survival under unfavorable conditions [17]. However, the stress response of other canonical nitrifiers is different from that observed in CMX bacteria. For example, a model AOA, that is, Nitrosopumilus maritimus SCM1, was adversely affected by NH3 starvation and Cu stress, leading to the downregulation of genes responsible for energy generation and biosynthesis [114]. These previous findings clearly imply that CMX bacteria have better stress adaptability than canonical nitrifiers.
Nonetheless, CMX Nitrospira seems to be a potential candidate in withstanding reactor conditions because of the adaptability of these bacteria. Resistance to physical and chemical stress is a key mechanism of the adaptability of CMX Nitrospira. It should be noted that the occurrence of some chemicals, such as ciprofloxacin, may significantly inhibit CMX Nitrospira and nitrogen removal [47]. Furthermore, in comparison with those in activated sludge, CMX Nitrospira in biofilm aggregates are expected to have higher operational flexibility, lower space requirements, and better resilience to environmental fluctuations [115], which has great implications for the real application of CMX-centered processes.
《5. Pollutant removal in WTPs and WWTPs 》
5. Pollutant removal in WTPs and WWTPs
《5.1. Biodegradation of micropollutants》
5.1. Biodegradation of micropollutants
Interestingly, some micropollutants can be exclusively degraded by CMX bacteria, which provides promising application of CMX in WTPs and WWTPs. The fate of micropollutants in the nitrifying environment was investigated recently [36], and it was found that Nitrospira inopinata possesses a high affinity for the NH3-degrading AMO system and degrades asulam, fenhexamid, mianserin, fantidine, and carbendazim in the presence of NH3. The presence of active genes responsible for the utilization of urea, amines and cyanates strongly suggests that CMX bacteria can potentially degrade diverse organic nitrogen compounds except for free NH3 [52]. It is noteworthy that increasing the dosage (0– 30 mg·L–1) of powdered activated carbon (PAC) to a nitrifying MBBR resulted in increased removal (up to 80%) of four different pharmaceuticals, where Nitrospira spp. were the dominant nitrifying species (8%–20%) in both the control and PAC-dosed reactors at all sampling times (79–164 d) [53]. However, the roles and mechanisms of CMX bacteria in the turnover of micropollutants are far from completely understood and deserve further study in the near future.
In WTPs or WWTPs, nitrifying bacteria normally play an important role in the degradation of different micropollutants, such as ibuprofen and naproxen [116], mianserin and ranitidine [117], sulfamethoxazole, erythromycin, roxithromycin and fluoxetine [118], and trichloroethane [119]. AMO could have a significant role in the hydroxylation of pharmaceuticals [118]. Enzyme-based studies also confirmed that AMOs can potentially oxidize a wide range of aliphatic and aromatic hydrocarbons [120] and micropollutants [121]. Importantly, other enzymatic systems in AMOs, such as hydroxylamine oxidoreductase (which catalyzes hydroxylamine to nitrite) and Nxr (which converts nitrite to nitrate), mediate oxidative biotransformation reactions and can play important roles in micropollutant removal [121]. The tolerance of pure-species cultures of ammonia oxidizers to heavy metals (e.g., Cu, Ni, Pb, and Cd) at varying concentrations (100–1000 μg·L–1) [122] implies that nitrifying populations seem to be potential candidates in the bioremediation of heavy metal-contaminated sites. To date, there is limited evidence for the successful applications of CMX bacteria either for the removal or biotransformation of micropollutants in engineered systems [20,36]. However, future studies are greatly warranted. On the one hand, more practical validation is needed to confirm the potential of CMX bacteria to degrade and/or transform micropollutants in different chemical groups. On the other hand, the ecotoxicity of micropollutants to CMX bacteria or the interaction between micropollutants and CMX bacteria should be investigated.
《5.2. Co-metabolic biotransformation》
5.2. Co-metabolic biotransformation
CMX bacteria have a propensity to exhibit co-metabolic biotransformation of micropollutants through deamination during simultaneous NH3 oxidation, such as the transformation of pharmaceuticals [54,55], artificial sweeteners (e.g., acesulfame, aspartame, cyclamate, saccharin, and sucralose) [56], sulfamethoxazole [57], and 2-chlorophenol [58]. In an aerobic granular reactor, the removal of selected micropollutants (e.g., bisphenol A, naproxen, irgarol, terbutryn, and iohexol) was particularly enhanced during simultaneous nitrification due to co-oxidation by AMOs [123]. Similarly, there was partial degradation of acidic pharmaceuticals (e.g., gemfibrozil, diclofenac, bezafibrate, and ketoprofen) and 17α-ethinylestradiol by nitrification, but the degradation rates of natural estrones (e.g., 17β-estradiol and estrone) were improved when nitrification was inhibited [124]. These previous attempts clearly imply that nitrifying enzymes (particularly AMOs) seem to be nonspecific [125] and could co-degrade several kinds of hydrocarbons during nitrifying reactions. It also indicates that the nitrifying community can be broadly applied to remove various micropollutants in WTPs or WWTPs. Certain micropollutants show either recalcitrance or inhibitory effects on the nitrifying process. For example, nitrifying activated sludge was found to be ineffective against diclofenac, diazepam, carbamazepine and trimethoprim [118]. Nonetheless, some emerging contaminants are toxic to bacteria, for example, azoles are known to inhibit nitrification [126]. The toxicity of azoles is attributed to the modification of active sites of nitrifying enzymes with substituent groups. Although the specific roles of CMX, AOA, AOB, and NOB in the co-metabolic biotransformation of micropollutants have not been well revealed, the co-metabolic capability of such functional bacteria will provide more alternatives for water or wastewater treatment.
It is widely acknowledged that micropollutant removal and cometabolic biotransformation can be considered effective features during the treatment of water and wastewater; fortunately, CMX Nitrospira has potential in these activities. Notably, an environmentally and economically sustainable WTP and WWTP have yet to emerge [127] because there is convincing evidence for the occurrence of micropollutants in treated water or wastewater [128–130]. On the other hand, co-metabolic biotransformation of organic micropollutants is ubiquitous in treatment facilities, such as activated sludge systems [131] and canonical nitrification systems [132,133]. Overall, CMX Nitrospira can aid in eliminating micropollutants via co-metabolic biotransformation.
《6. Knowledge gaps》
6. Knowledge gaps
The shaping of CMX for sustainable water or wastewater treatment depends on many variables and requires comprehensive studies. These include:
(1) Niche preferences under different trophic levels and operational conditions of reactors should be clarified in future studies. Although niche preferences of CMX have been partially revealed [13,16,30], knowledge regarding resource utilization, niche specialization and niche overlaps of different clades of CMX bacteria is still limited. In particular, niche overlap other than competitive exclusion tends to occur when two species possess similar competitive abilities in source utilization [134].
(2) Switches between the modes of NH3 and NO2 oxidation in CMX bacteria remain unknown. Note that environmental factors are the key drivers of such switches [135]. It should be kept in mind that CMX bacteria harbor higher nitrification rates in a reactor dosed with urea than NO2– as a sole nitrogen source [102]. However, it is difficult to clarify the factors determining nitrogen preference [10].
(3) Reactor-centered studies focusing on the determination of boundary conditions should be extensively conducted, which can help to realize early large-scale application of CMX bacteria. Interactions of CMX bacteria with other bacterial groups are still limited in both natural and engineered systems. Therefore, future exploration of bacterial interactions between CMX and other bacteria will definitely facilitate the evaluation of reactor adaptability of CMX bacteria.
(4) Deep insights into the stress response of CMX bacteria remain to be explored. Stress-related genetic elements in CMX bacteria can help in understanding the mechanisms of bacterial persistence in nonhost environments [136], which deserve further investigation. It should be noted that water or wastewater has unique and distinct microbial and chemical compositions, which will also impose stresses on CMX.
(5) The role of CMX in the turnover of micropollutants is not completely known. This requires in-depth analysis of many issues, such as the abundance of CMX in engineered ecosystems, potential of CMX bacteria to degrade micropollutants, and factors influencing micropollutant removal by CMX bacteria. Addressing these issues could help us to design an efficient process to remove micropollutants using CMX bacteria in the future.
(6) Currently, we know that CMX Nitrospira is ahead compared with canonical nitrifiers by exhibiting higher DO affinity and thriving under low DO concentrations, but it is doubtful whether CMX Nitrospira is truly an effective energy saver while removing nitrogen. It is also worth focusing on manipulations of enzyme activities to reduce the energy demand of CMX Nitrospira.
《7. Conclusions》
7. Conclusions
CMX bacteria are ubiquitously distributed in WTPs and WWTPs. Importantly, CMX bacteria have several competitive advantages over coresident canonical nitrifiers, for example, the ability to grow at low DO levels, rapid niche divergence with horizontal gene transfer, urea utilization with high affinity, and efficient nitrogen removal at low DO and high SRT. Furthermore, CMX bacteria can endure long-term stress induced by diverse chemicals and can also co-metabolize certain pollutants (e.g., pharmaceuticals and personal care products (PPCPs)) during simultaneous NH3 oxidation. Although CMX bacteria coexist with various canonical nitrifiers, each clade of CMX bacteria has its own coresident bacterial community. It can be concluded that the use of CMX bacteria in WTPs or WWTPs for nitrogen and micropollutant removal is practically possible. However, future studies are largely needed for better design and operation of CMX bacteria-based reactors.
《Acknowledgments 》
Acknowledgments
This work was supported by the National Natural Science Foundation of China (52000185 and 51878675), Key Research and Development Program of Shandong Province (2019JZZY020308), and China Postdoctoral Science Foundation, China (2017M612807).
《Compliance with ethical guidelines 》
Compliance with ethical guidelines
Naga Raju Maddela, Zhihao Gan, Yabing Meng, Fuqiang Fan, and Fangang Meng declare that they have no conflicts of interest or financial conflicts to disclose.











 京公网安备 11010502051620号
京公网安备 11010502051620号




