

《1 工程研究前沿》
1 工程研究前沿
《1.1 Top 11 工程研究前沿发展态势》
1.1 Top 11 工程研究前沿发展态势
农业领域 Top 11 工程研究前沿主要有:① 有关农业动植物的分子生物学机制和机理的研究,如重要动物病原的免疫抑制与逃逸机制、土壤高效固碳与调控机制、木材形成的分子生物学机制、养殖环境 – 畜禽 – 肠道微生物 – 营养素代谢互作网络机制;② 一如既往的分子育种,如水产动物多倍体育种、畜禽多基因聚合育种;③ 关于提升动植物产品产量及绿色生产相关研究,如园艺作物产品器官发育与品质调控、作物绿色栽培技术、植物抗病小体的发现、作物从头驯化及野生种质资源开发利用、粮食安全对气候变化的响应。农业领域工程研究前沿的核心论文数区间为 7~143 篇,平均为 59 篇,与往年相近;篇均被引频次区间为 5.06~178.69 次,平均约为 59.58 次;核心论文出版年度以 2018年和 2019 年为主,其中“重要动物病原的免疫抑制与逃逸机制”和“作物从头驯化及野生种质资源开发利用”的核心论文出版以近 3 年为主,且呈上升趋势(表 1.1.1 和表 1.1.2)。
(1) 作物从头驯化及野生种质资源开发利用
作物驯化是将野生植物驯化繁殖为栽培作物,其在人类农耕文明的起源和演变过程中发挥了重要作用 , 推动了人类文明的持续发展和社会的快速进步。具体来看,驯化过程是以野生种质为起始材料, 通过历史上长期的人工选择有目的地筛选有益农艺性状,如落粒少、少分枝、直立生长、芒短、种子大、易收获等。究其本质,驯化过程是聚合和保留基因组中优异遗传变异的过程。然而,驯化过程历时漫长,同时对性状的选择较为朴素盲目,因此筛选驯化优异性状的同时不可避免地使驯化群体遗传多样性降低。相较于现代栽培种,野生近缘种往往在特定性状方面具有优势,包括具有更强的抗性和更广的环境适应性,生物量与经济产量会相对较高。
《表 1.1.1》
表 1.1.1 农业领域 Top 11 工程研究前沿
| 序号 | 工程研究前沿 | 核心论文数 | 被引频次 | 篇均被引频次 | 平均出版年 |
| 1 | 作物从头驯化及野生种质资源开发利用 | 74 | 2 345 | 31.69 | 2019.4 |
| 2 | 重要动物病原的免疫抑制与逃逸机制 | 45 | 7 556 | 167.91 | 2020.2 |
| 3 | 土壤高效固碳与调控机制 | 83 | 14 831 | 178.69 | 2018.9 |
| 4 | 园艺作物产品器官发育与品质调控 | 61 | 588 | 9.64 | 2018.9 |
| 5 | 水产动物多倍体育种 | 18 | 91 | 5.06 | 2019.2 |
| 6 | 作物绿色栽培技术 | 98 | 4 868 | 49.67 | 2017.6 |
| 7 | 畜禽多基因聚合育种 | 50 | 569 | 11.38 | 2019.2 |
| 8 | 粮食安全对气候变化的响应 | 143 | 11 032 | 77.15 | 2017.8 |
| 9 | 木材形成的分子生物学机制 | 7 | 167 | 23.86 | 2017.7 |
| 10 | 植物抗病小体的发现 | 34 | 800 | 23.53 | 2020.2 |
| 11 | 养殖环境 – 畜禽 – 肠道微生物 – 营养素代谢互作网络机制 | 37 | 2 842 | 76.81 | 2017.3 |
《表 1.1.2》
表 1.1.2 农业领域 Top 11 工程研究前沿核心论文逐年发表数
| 序号 | 工程研究前沿 | 2016 | 2017 | 2018 | 2019 | 2020 | 2021 |
| 1 | 作物从头驯化及野生种质资源开发利用 | 4 | 10 | 6 | 12 | 13 | 29 |
| 2 | 重要动物病原的免疫抑制与逃逸机制 | 0 | 0 | 4 | 5 | 15 | 21 |
| 3 | 土壤高效固碳与调控机制 | 9 | 11 | 7 | 25 | 15 | 16 |
| 4 | 园艺作物产品器官发育与品质调控 | 4 | 10 | 14 | 9 | 7 | 17 |
| 5 | 水产动物多倍体育种 | 2 | 2 | 1 | 3 | 5 | 5 |
| 6 | 作物绿色栽培技术 | 29 | 17 | 26 | 21 | 5 | 0 |
| 7 | 畜禽多基因聚合育种 | 0 | 7 | 11 | 12 | 6 | 14 |
| 8 | 粮食安全对气候变化的响应 | 32 | 32 | 34 | 25 | 16 | 4 |
| 9 | 木材形成的分子生物学机制 | 2 | 2 | 1 | 0 | 2 | 0 |
| 10 | 植物抗病小体的发现 | 0 | 1 | 0 | 6 | 11 | 16 |
| 11 | 养殖环境 – 畜禽 – 肠道微生物 – 营养素代谢互作网络机制 | 15 | 6 | 9 | 4 | 3 | 0 |
如今随着生物技术特别是基因编辑技术的发展,从头驯化开始被提出作为作物育种的一种新策略。选择野生或半野生植物物种作为优良基础材料,通过现代育种技术手段快速导入驯化目标性状,短周期内实现培育突破性新作物物种,以重新挖掘利用在传统作物驯化过程中丢失的优异野生种质资源,对适应当前环境变化大趋势下的农业可持续发展有重要意义。
(2) 重要动物病原的免疫抑制和逃逸机制
我国是世界畜禽养殖和畜禽类动物源食品消费大国,近年来畜牧业集约化和规模化养殖水平不断提高,但疫病问题尤其是一些重大动物疫病及人兽共患病一直是制约养殖业健康发展和社会公共卫生安全的主要因素。如何有效防控这些重要动物病原一直是兽医学、医学和生物安全等领域的关注焦点和研究热点。阻碍疫病防控的主要原因之一就是这些重要动物病原在长期进化中产生不同机制的免疫抑制和免疫逃逸。机体抵御病原感染主要依靠机体天然免疫和获得性免疫应答。一些重要病原能够通过逃逸宿主识别和抑制关键抗病毒天然免疫应答, 导致病原能够有效突破机体的免疫屏障建立感染; 同时,病原通过抑制宿主诱导获得性应答产生,通过突变或重组等机制产生新的变异体或毒株,逃逸机体产生的中和抗体或 T 细胞免疫,从而免于被机体清除,建立有效感染。揭示病原抑制机体免疫功能和逃避免疫系统的关键机制是有效防控重要动物病原的前提,也是当前研究的难点和热点,相关研究成果必将为防控疫病的发生与流行提供重要指导和技术支撑。
(3) 土壤高效固碳与调控机制
全球土壤有机碳总量为 1 500~2 000 Pg,相当于大气中碳总量的 2~3 倍。农业土壤碳固定占自然气候解决方案总潜力的 25%。因此,促进农业土壤高效固碳对于保障粮食安全、减缓气候变化以及推进农业绿色发展具有重要意义。土壤高效固碳与调控的核心是深入认识土壤有机质的形成、周转和稳定等关键过程,建立高效固碳、减缓其矿化分解的调控机制。
土壤是一个多种物质并存、多种过程同时发生、多种因素共同影响的开放的复杂系统。目前土壤高效固碳机理还存在很多不清楚的地方。因此未来需要强化对其固碳过程和调控机制的深入研究。具体来看,解析土壤有机质的形成过程、赋存形态和稳定机理等问题是该方向重要的研究热点和前沿,未来应精确区分不同有机组分碳源、探究植物残体向土壤有机质转化的微生物作用机制、加快基于多因素协同调控的有机质稳定性研究、探索土壤碳平衡机理及植物残体输入阈值等,强化对土壤固碳本质和调控机制的研究。此外,创新高效固碳保肥新型碳基材料,提高土壤固碳效率,集成创新土壤固碳、耕地保育及产能提升关键技术及技术模式,推动大面积农业土壤固碳落地应用已成为该领域的重要应用研究前沿。
(4) 园艺作物产品器官发育与品质调控
园艺作物(蔬菜、果树、花卉)产品器官多样, 包括根及变态根、茎及变态茎、叶及叶球、花及花薹、果实及种子等,是园艺作物产量构成的基础。园艺产品品质包括营养品质(如碳水化合物、脂类、蛋白质、维生素、矿物质、微量元素等营养要素)、感官品质(包括产品的外观、质地、适口性等, 如大小、形状、颜色、光泽、汁液、硬度、缺陷、新鲜度等)、卫生品质(包括果蔬表面的清洁程度, 果蔬组织中的重金属含量、农药残留量及其他限制性物质如亚硝酸盐含量等)、商品化处理品质(如易清洗等)等复杂性状。随着园艺产业的快速发展, 在不断提高作物产量的同时,产品品质性状变化、健康成分含量和园艺产品质量安全问题受到了密切关注。近年来,我国在园艺作物果形、色泽、营养品质、风味和苦味物质形成与调控机理方面已开展了一些研究,特别是运用基因组、转录组和代谢组等手段,通过园艺作物器官发育与产品品质相关物质代谢基因及其调控基因的挖掘、分析,对产量与品质等重要农艺性状分子机制和调控网络的了解也越来越多,但多数决定园艺作物器官发育与产品品质的代谢物质还不清晰,对其分子机制和代谢调控机理的研究更少。因此,未来应该在现有基础上,从转运蛋白、转录调控因子、表观修饰因子以及非编码 RNA 等方面开展不同层级的基因表达级联调控机制及其调控网络解析, 探明园艺作物产品器官发育与品质形成的调控网络和信号传导机制,为园艺产业提质增效提供科学依据。
(5) 水产动物多倍体育种
多倍体生物是具有 3 套或 3 套以上完整染色体组的生物。在植物和动物中,多倍体广泛存在。在长期的进化过程中,多倍化是物种发生的重要原动力之一。随着基因组和系统演化研究的深入,多项研究结果支持大部分物种在演化过程中经历了多倍化事件。一旦多倍化发生,其稳定性则依赖于基因组快速重组和基因表达调控的变化。可育多倍体的形成不仅促进了物种间的遗传物质交流,丰富了物种多样性,而且为多倍体育种奠定了基础。通过远缘杂交、物理诱导、化学诱导等技术可以获得水产动物多倍体。其中远缘杂交可获得大规模的多倍体可育品系。目前,研究人员基于水产动物多倍体育种技术已经获得了多个鱼类、虾蟹、贝类等水产动物新种质,并开展了系统研究。水产动物多倍体育种的研究不仅具有重要的理论意义 , 而且具有重要的应用价值。
(6) 作物绿色栽培技术
绿色栽培是研究作物生长发育规律及其与外界环境条件的关系,并在此基础上通过栽培措施达到高产、优质、高效、生态、安全的一门应用科学领域。我国水稻、小麦、玉米三大作物单产显著高于世界平均水平,粮食总产量连创新高,为保障国家粮食安全做出了巨大贡献。但应清醒认识到,我国粮食作物种植方式多元且趋向粗放、机械化栽培技术不完善、工程技术集成度不高、水肥药投入多、农艺农机农智绿色低碳融合不足的问题突出。作物绿色栽培已成为当前亟待解决的热点和难点问题, 其核心科学问题主要有:① 大田作物优质高产绿色高效协同提升机理与技术;② 大田作物优质高产(超高产)高效协同规律与可复制栽培模式;③ 大田作物健康抗逆固碳减排节能绿色栽培技术;④ 大田作物农艺农机农智融合关键技术;⑤ 作物全程“无人化”绿色栽培模式与技术。通过作物栽培学的自主创新及与生理学、环境生态学、信息学、机械工程学等前沿科学的交叉融合,创建中国特色作物优质高产绿色高效栽培技术新理论,创立面向规模化绿色生产的农艺农机农智协同的“无人化” 栽培技术体系,实现大田作物产量与品质、生产效率与生产效益的协同提升,作物综合生产能力提升10%~30%,绿色高产栽培居世界领先水平,有效推进大田作物生产现代化。
(7) 畜禽多基因聚合育种
畜禽多基因聚合育种是畜禽分子育种的主要方法之一,是指将分散在不同品种或品系中的优良个体的优良基因聚合到同一个体的基因组中,从而获得具备特定性状的新品种(系)。目前,实现多基因聚合主要有两种途径。一是在确定与优异性状相关的分子标记的基础上,通过杂交、回交和分子标记辅助选择技术,在后代中选择多个优良基因聚合到一起的个体。分子标记辅助选择所获得的与目标数量性状基因紧密连锁的分子标记,其结果可靠性强,且不受等位基因显隐性关系及环境的影响,在动物育种中的应用加快了遗传进展,缩短了育种周期。但是该方法选择的个体后代中聚合的基因可能重新分离,造成目标性状不稳定。近年来,随着转基因技术和基因编辑技术的不断发展,通过基因修饰技术实现多基因聚合已经成为畜禽多基因聚合育种的研究前沿。但是,在动物基因组中同时实现多基因编辑仍然有多重技术难点。一是与畜禽重要性状相关的功能基因没有完全解析清楚,基因组操作的靶点仍不明确。二是在动物基因组中同时进行基因编辑的数量有限,同时编辑几十、上百个位点难以实现。我国拥有丰富的畜禽遗传资源,各种表型和基因型数据库不断丰富,多种性状相关的重要功能基因和调控序列挖掘不断深入,同时,随着干细胞育种技术以及基因编辑技术的效率、准确性和安全性的不断提升,利用干细胞体外长期传代培养的特性,能够进行多次基因编辑操作,有望解决基因编辑聚合育种的技术难题。
(8) 粮食安全对气候变化的响应
全球气候变化是指在全球范围内气候平均状态统计学意义上的巨大改变或者持续较长一段时间的气候变动。粮食生产对于气候变化的响应具有高度的敏感性,以气候变暖为主要特征的全球气候变化给全球粮食安全带来了严重影响。随着全球气候变暖,地表温度上升会增加农作物的呼吸消耗,影响光合作用的进行和农作物生长发育。在全球范围内, 玉米产量年际波动中有 18% 是气候变化的结果。气候变异对大豆和小麦产量波动的影响分别占其综合影响的 7% 和 6%。相关研究表明,到 2050 年, 全球气候变暖可能导致世界粮食产量减少 18%。目前,气候变化已对人类基本生存和社会经济稳定构成重大威胁,阐明气候变化驱动机制,以及对作物生产影响的规律已成为当前亟待解决的热点和难点问题,其核心科学问题主要有:① 气候变化核心驱动因素识别;② 农田生态系统水通量及生产力对全球变化的响应机制;③ 气候变化和人类活动作用下作物灾损变化机制。随着人工智能和大数据技术的发展,可以从全球尺度辨识气候变化的核心驱动因素,并从自然、经济、政策等多维度分析全球作物产量变化规律,为应对全球气候变化对粮食产量的消极影响提供科学决策依据,为构建全球人类命运共同体提供保障。
(9) 木材形成的分子生物学机制
木材形成从树木维管形成层细胞分裂产生新的木质部前体细胞开始,经过细胞的分化、伸长和扩展、次生细胞壁合成与沉积,细胞程序性死亡等阶段,形成细胞壁特异加厚的木材组织。目前主要的研究方向有:① 次生细胞壁主要组分的合成机理;② 生长素、细胞分裂素、短肽和转录因子等对维管形成层细胞发生、增殖以及木质部细胞分化的调控;③ 形成层干细胞维持,分化以及协同调控木质部发育与环境适应性机制;④ 蛋白翻译后修饰和表观遗传修饰在形成层活性和木材发育中的调控作用;⑤ 应拉木形成的分子机制;⑥ 生长与木材品质性状表观数量性状基因座(quantitative trait locus,QTL)定位研究;⑦ 重要材性性状的全基因组关联分析。其发展趋势主要有:① 对木材形成研究更注重应用先进的研究方法、技术和分析仪器,如时空转录组、空间代谢组、单分子成像、原子力显微镜等;② 建立精细的多层级转录调控网络以及不同层级的反馈调节机制;③ 重要木材品质和材性性状的挖掘鉴定及层级调控机制的解析;④ 利用新型传感器技术进行材质信息的采集和分析;⑤ 关键调控蛋白的晶体结构解析和蛋白互作面位点鉴定;⑥ 基因编辑技术创制品质优良的林木新品种;⑦ 开展林木分子设计育种,培育速生、优质、高产林木新品种。
(10) 植物抗病小体的发现
病菌在侵染植物时往往会分泌多种多样的致病性蛋白去干扰植物的生理活动,这些蛋白称为效应蛋白。植物中存在一类以 NB-LRR(nucleotide- binding site, leucine-rich repeat)结构为基础的抗病蛋白,这些蛋白可以直接或间接识别病菌效应蛋白, 进而启动免疫反应,产生抗病性。自从 Harold Flor 于 1956 年提出“基因对基因”(gene for gene)理论以来,人们对于植物抗病性的理解就是 NB-LRR 基因编码的抗病蛋白对应于病菌中的某个效应蛋白,产生识别启动免疫。这一识别产生的结果通常是强烈的细胞超敏反应而导致的细胞死亡,进而可以限制病菌的入侵,也是 NB-LRR 介导植物抗病性的基础。尽管自 20 世纪 90 年代以来,基于“基因对基因”理论克隆了很多病菌中的效应蛋白以及对应的植物中的抗病 NB-LRR 蛋白,然而很长一段时间,对于 NB-LRR 蛋白是如何识别效应蛋白并启动免疫反应(如超敏细胞死亡)的,一直没有获得突破,其中一个重要的难点是无法获得完整的NB-LRR 蛋白结晶,因而无法从结构上给予揭示。2019 年,中国科学院遗传与发育生物学研究所与清华大学合作发现了黄单胞菌效应蛋白 AvrAC 激活的 NB-LRR 与相关蛋白形成的复合体,该复合体是一个五聚体复合物。激活后的五聚体复合物靶向细胞膜,形成钙离子通道,造成钙离子内流,诱发活性氧在细胞内大量积累,导致细胞发生超敏反应死亡。由于该复合物的作用机制类似于 NAIP2- NLRC4(NB-LRR 蛋白)形成的抗病炎症小体,因而称之为植物的抗病小体。2020 年,德国和美国的科学家又发现了相似的四聚抗病小体复合物。这些工作揭开了 NB-LRR 蛋白识别病菌效应蛋白并启动免疫反应的分子机制,是植物病理学研究中的里程碑事件。
(11) 养殖环境 – 畜禽 – 肠道微生物 – 营养素代谢互作网络机制
畜牧生产环境污染问题严重压缩畜牧业发展空间,新型生产生态系统尚处于探索阶段。原国家环保总局调查显示:我国每年产生的畜禽粪便量约为 19 亿 t,是工业固体废弃物的 2.4 倍;我国畜牧业化学需氧量(chemical oxygen demand, COD)年排放 1 200 多万吨,占农业排放总量的95.8%、占全国 COD 排放总量的 41.9%,超过工业污染。针对养殖效率和环境污染以及畜禽舒适度与畜舍环境参数控制问题,需重点解决改善饲料品质、提高饲料营养价值和利用效率、降低加工成本、精准饲料配制及环境友好型饲料生产、实现精准饲养的自动化和智能化设备研发等重大技术瓶颈。发展饲料养分高效利用的体外预消化新技术;研究畜禽饲料营养高效利用与转化机理, 针对不同饲料原料,利用微生物培养组学技术、微生物改造工程技术、蛋白质工程技术、合成生物学技术,发展微生物细胞工厂,创制饲料高效利用、抗营养因子及毒素消除的体外预消化添加剂;基于菌酶协同发酵工艺路线,以酸溶蛋白含量和抗营养组分及毒素的降解率为目标,建立关键工艺参数、安全评价和质量控制体系。有效提高饲料养分的利用率,减少饲料蛋白的添加,减少饲料抗营养因子及毒性成分,促进饲料的消化吸收,有效减少环境污染。
《1.2 Top 3 工程研究前沿重点解读》
1.2 Top 3 工程研究前沿重点解读
1.2.1 作物从头驯化及野生种质资源开发利用
(1) 作物从头驯化及野生种质资源开发利用研究的重要意义
作物驯化是将野生植物驯化繁殖为栽培作物, 其在人类农耕文明的起源和演变过程中发挥了重要作用 , 推动了人类文明的持续发展和社会的快速进步。具体来看,驯化过程是以野生种质为起始材料, 通过历史上长期的人工选择有目的地筛选有益农艺性状,如落粒少、少分枝、直立生长、芒短、种子大、易收获等。究其本质,驯化过程是聚合和保留基因组中优异遗传变异的过程。然而,驯化过程历时漫长,同时对性状的选择较为朴素盲目,因此筛选驯化优异性状的同时不可避免地使驯化群体遗传多样性降低。相较于现代栽培种,野生近缘种往往在特定性状方面具有优势,包括具有更强的抗性和更广的环境适应性,生物量与经济产量会相对较高。如今随着生物技术特别是基因编辑技术的发展,从头驯化开始被提出作为作物育种的一种新策略。选择野生或半野生植物物种作为优良基础材料,通过现代育种技术手段快速导入驯化目标性状,短周期内实现培育突破性新作物物种,以重新挖掘利用在传统作物驯化过程中丢失的优异野生种质资源,对适应当前环境变化大趋势下的农业可持续发展具有重要意义。
(2) 作物从头驯化及野生种质资源开发利用研究现状
现代作物驯化伴随的负向效应。野生物种驯化后的作物适应各种气候和土壤条件,使得早期种植业可以扩展到更大的区域,而随后的作物育种催生了更高的粮食产量并促进了人口增长和人类文明繁衍。驯化与育种过程相似,都是选择最佳表型,即基因型与环境的最佳组合。很多作物驯化前后的性状改变有一定的共通规律,包括株型直立紧凑、穗型增大、籽粒增多、落粒性丧失、休眠性降低、开花时间以及种子色素沉积的改变。新的基因型由突变产生并表现出新的性状,而不同基因型依赖交换重组实现不同优异性状在单株植物中的聚合。然而,基因型突变和交换重组都是偶然发生的不定向过程,驯化者或育种者无法准确预测。同时,基因组中的自发突变频率极低,驯化或育种过程中的外源基因组片段渗入也可能抑制重组,导致有害等位基因在这些区域固定,而造成连锁累赘、优异等位变异无法聚合。例如在番茄中的根结线虫抗性(由Mi-1 基因控制)和黄叶卷曲病毒抗性(由 Ty-1 基因控制)均位于 6 号染色体上一个交换重组抑制区内,遗传距离很近,同时聚合这两个抗性性状必须借助大量、长期的遗传筛选。另外,驯化和早期育种过程将目标过多聚焦于作物产量提升 , 基因组中大量的优质基因因未被选择而丢失 , 造成“驯化瓶颈”;对控制某一特定目标性状基因区域的选择, 使该区域和其相邻区间的遗传多样性显著降低,出现“选择性清除”,致使现有栽培作物遗传多样性降低,抗逆和某些品质营养性状较差,品种同质化严重。
驯化和早期育种当时都是凭经验进行的,对其背后的生物机制了解甚少。随着近代驯化群体遗传学的发展,尤其是新兴的泛基因组测序的兴起,不断深入了人们对作物驯化遗传机理的认识。不同作物经历成百上千年的长期驯化过程,然而对 200 余种作物的驯化综合性状分析表明,84% 的作物仅有2~5 个主要性状为主要的驯化性状。更重要的是, 以玉米为代表的驯化遗传学分析结果证实,作物驯化往往由少数几个关键基因控制。通过对作物驯化遗传机理和基因组学基础的深入解析,结合快速发展的基因组编辑技术,通过对优异性状的作物野生近缘种进行从头驯化或半驯化作物的再驯化的时代已经来临。2018 年,中外两组科研团队同时报道了野生醋栗番茄的从头驯化,保持野生番茄优异耐盐碱和抗病能力的前提下,培育出株型优异、产量品质协同提升的新型番茄。同年,美国科学家也成功对半驯化小众作物灯笼果进行了再驯化 , 快速实现了产量和综合农艺性状的突破性提高。2021 年中国科学家团队首次实现了快速从头驯化异源四倍体野生稻,为最终培育出产量高、环境适应能力强的新型水稻作物开辟了全新的育种方向。
(3) 未来研究方向与创新点
为进一步挖掘野生种质资源,拓展从头驯化的应用范围,作物从头驯化的技术潜力需从以下几个方向进一步提升:① 深化主要作物驯化遗传机理的解析,挖掘物种间共性或物种特异的关键驯化性状相关基因,为从头驯化提供基因基础;② 野生种质资源的收集与遗传鉴定,筛选具备部分优异农艺性状的野生种或半驯化物种,为从头驯化提供种质基础;③ 野生物种中高效快速的遗传转化体系的建立和持续优化,打破多数野生种难以遗传转化的瓶颈,为从头驯化提供技术先决条件;④ 野生种中高效基因编辑系统的建立和持续优化,实现野生种中精准调控的基因敲除、单碱基编辑、引导编辑、表达量激活或抑制编辑等;⑤ 高通量驯化表型鉴定,实现大规模转化体的优异性状精准高效筛选。
在“作物从头驯化及野生种质资源开发利用” 工程研究研究中,核心论文数排名前三位的国家分别是中国(占 51.35%)、美国(占 37.84%)和德国(占 13.51%)(表 1.2.1)。该领域核心论文篇均被引频次分布在 12.00~127.00,其中以色列和沙特阿拉伯篇均被引频次均超过了 100。该领域核心论文的主要产出机构(表 1.2.2)方面,中国科学院、
《表 1.2.1》
表 1.2.1 “作物从头驯化及野生种质资源开发利用”工程研究前沿中核心论文的主要产出国家
| 序号 | 国家 | 核心论文数 | 论文比例 /% | 被引频次 | 篇均被引频次 | 平均出版年 |
| 1 | 中国 | 38 | 51.35 | 1 189 | 31.29 | 2019.8 |
| 2 | 美国 | 28 | 37.84 | 1 486 | 53.07 | 2019.2 |
| 3 | 德国 | 10 | 13.51 | 633 | 63.3 | 2019.5 |
| 4 | 澳大利亚 | 9 | 12.16 | 844 | 93.78 | 2019 |
| 5 | 英国 | 7 | 9.46 | 456 | 65.14 | 2019.1 |
| 6 | 法国 | 6 | 8.11 | 197 | 32.83 | 2018.8 |
| 7 | 日本 | 6 | 8.11 | 72 | 12 | 2020.3 |
| 8 | 巴西 | 5 | 6.76 | 332 | 66.4 | 2019.2 |
| 9 | 以色列 | 3 | 4.05 | 381 | 127 | 2019 |
| 10 | 沙特阿拉伯 | 3 | 4.05 | 309 | 103 | 2020.3 |
《表 1.2.2》
表 1.2.2 “作物从头驯化及野生种质资源开发利用”工程研究前沿中核心论文的主要产出机构
| 序号 | 机构 | 核心论文数 | 论文比例 /% | 被引频次 | 篇均被引频次 | 平均出版年 |
| 1 | 中国科学院 | 13 | 17.57 | 779 | 59.92 | 2019.8 |
| 2 | 中国农业科学院 | 8 | 10.81 | 197 | 24.62 | 2020.8 |
| 3 | 华中农业大学 | 7 | 9.46 | 181 | 25.86 | 2020 |
| 4 | 美国农业部农业研究局 | 4 | 5.41 | 359 | 89.75 | 2019 |
| 5 | 浙江大学 | 4 | 5.41 | 336 | 84 | 2019.8 |
| 6 | 明尼苏达大学 | 4 | 5.41 | 329 | 82.25 | 2018.2 |
| 7 | 圣保罗大学 | 4 | 5.41 | 326 | 81.5 | 2019.2 |
| 8 | 维索萨联邦大学 | 3 | 4.05 | 300 | 100 | 2020 |
| 9 | 亚利桑那大学 | 3 | 4.05 | 198 | 66 | 2019.7 |
| 10 | 图卢兹大学 | 3 | 4.05 | 143 | 47.67 | 2019.7 |
中国农业科学院和华中农业大学产出的核心论文数及被引频次较高。主要国家间的合作网络(图 1.2.1) 方面,国家间的研究合作较为普遍,以中国、美国、德国之间合作相对更紧密。主要机构间的合作网络(图 1.2.2)方面,各机构间均存在一定的合作关系。施引核心论文的主要产出国家是中国、美国和澳大利亚,中国占比超过 1/3,美国占比为 20% 以上(表1.2.3)。施引核心论文的主要产出机构(表 1.2.4) 方面,中国农业科学院、中国科学院和美国农业部农业研究局的施引核心论文数排在前三位。图 1.2.3 为“作物从头驯化及野生种质资源开发利用”工程研究前沿的发展路线。
1.2.2 重要动物病原的免疫抑制和逃逸机制
我国是世界养殖大国和动物源食品消费大国, 生猪存栏占世界总量的 50%,年出栏约 7 亿头,养禽 140 亿羽,肉禽产量世界第一,全国畜牧业总产值超过 3.2 万亿元,占农业总产值比例接近 30%, 带动上下游相关产业产值在 3 万亿元以上。然而, 我国畜牧业的生产水平远低于世界平均水平,制约其发展的其主要瓶颈因素是疫病问题。非洲猪瘟、小反刍兽疫等动物疫病的不断发生给我国畜牧业造成了巨大的经济损失,严重影响畜产品的产量和质量,如 2018 年在我国首次暴发的非洲猪瘟,给我国养猪业造成毁灭性的破坏,2019 年生猪存栏量
《图 1.2.1》
图 1.2.1 “作物从头驯化及野生种质资源开发利用”工程研究前沿主要国家间的合作网络
《图 1.2.2》
图 1.2.2 “作物从头驯化及野生种质资源开发利用”工程研究前沿主要机构间的合作网络
《表 1.2.3》
表 1.2.3 “作物从头驯化及野生种质资源开发利用”工程研究前沿中施引核心论文的主要产出国家
| 序号 | 国家 | 施引核心论文数 | 施引核心论文比例 /% | 平均施引年 |
| 1 | 中国 | 863 | 35.81 | 2020.4 |
| 2 | 美国 | 499 | 20.71 | 2020.2 |
| 3 | 澳大利亚 | 164 | 6.8 | 2020.2 |
| 4 | 印度 | 160 | 6.64 | 2020.2 |
| 5 | 德国 | 159 | 6.6 | 2020.3 |
| 6 | 英国 | 142 | 5.89 | 2020.1 |
| 7 | 法国 | 119 | 4.94 | 2020 |
| 8 | 意大利 | 92 | 3.82 | 2020.3 |
| 9 | 日本 | 81 | 3.36 | 2020.1 |
| 10 | 西班牙 | 66 | 2.74 | 2020.2 |
《表 1.2.4》
表 1.2.4 “作物从头驯化及野生种质资源开发利用”工程研究前沿中施引核心论文的主要产出机构
| 序号 | 机构 | 施引核心论文数 | 施引核心论文比例 /% | 平均施引年 |
| 1 | 中国农业科学院 | 207 | 23.44 | 2020.5 |
| 2 | 中国科学院 | 129 | 14.61 | 2020.3 |
| 3 | 美国农业部农业研究局 | 91 | 10.31 | 2020.2 |
| 4 | 华中农业大学 | 83 | 9.4 | 2020.6 |
| 5 | 南京农业大学 | 68 | 7.7 | 2020.4 |
| 6 | 郑州大学 | 67 | 7.59 | 2020.7 |
| 7 | 中国农业大学 | 55 | 6.23 | 2020.6 |
| 8 | 浙江大学 | 55 | 6.23 | 2020.7 |
| 9 | 西澳大学 | 45 | 5.1 | 2020.2 |
| 10 | 昆士兰大学 | 43 | 4.87 | 2019.9 |
《图 1.2.3》
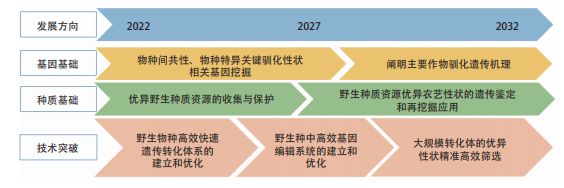
图 1.2.3 “作物从头驯化及野生种质资源开发利用”工程研究前沿的发展路线
下降 60%,有些地方甚至超过 70%。猪肉价格从每千克 22~24 元飙升到 60 多元。此外,布病、结核病、鼠疫、流感、狂犬病、埃博拉和新型冠状病毒肺炎等动物源性人兽共患病的流行对公共卫生安全造成严重威胁,如当前仍在肆虐全球的新型冠状病毒肺炎已导致 600 余万人的死亡。如何有效防控动物疫病和人兽共患病的发生与流行一直是兽医学、医学和生物安全领域关注的焦点和研究的热点。
然而,制约动物疫病及人兽共患病防控研究取得突破的关键原因就是病原的免疫抑制和免疫逃逸。机体抵御病原感染主要依靠机体免疫系统。免疫逃逸可导致病原能够有效突破机体的免疫屏障或疫苗诱导的免疫应答,建立有效感染。而免疫抑制可促进病原感染或者建立持续感染。例如流感病毒和冠状病毒经过长期演化都能够通过多种机制逃逸宿主的天然免疫应答建立感染,并不断产生新的变异毒株逃逸现有疫苗的保护。因此, 揭示病原抑制机体免疫功能和逃避免疫系统的机制是有效防控这些重要动物病原的前提,也是当前研究的热点和难点。
宿主免疫系统是抵御病原感染的关键力量,包括天然免疫和获得性免疫。宿主通过模式识别受体(pattern recognition receptor,PRR)监测到病原感染后最先诱导天然免疫应答,天然免疫应答是抵抗病原感染的第一道防线,而很多病原与其宿主在长期共同进化过程中获得能够逃避或主动抑制宿主免疫的能力。PRR 是抗感染免疫的关键成分,可检测病毒病原体的保守分子特征并启动天然免疫应答。因此,很多病原都存在免疫逃避或抑制的能力,其分子机制包括:①隔离或修饰病毒 RNA 或 DNA 核酸配体以逃逸细胞内 PRR 的识别和激活,如流感病毒的非结构蛋白 1(NS1)和牛痘病毒的 E3 蛋白通过与病毒 dsRNA 结合来避免被 RNA 感应器RIG-I 识别;②操纵 PRR 蛋白的翻译后修饰使其失去功能或降级,如冠状病毒的木瓜蛋白酶样蛋白酶(PLP)和口蹄疫病毒(FMDV)的先导蛋白酶(Lpro)可以切割或降解天然免疫中关键蛋白RLR 和 MAVS,以逃逸天然免疫应答。此外,许多病毒还通过靶向天然免疫应答共享的一些关键下游分子来抑制先天免疫反应,如 TBK1、IRF3、IRF7 和NF-κB,或者阻断 IFNα/β 受体的信号传导。
一些重要病原在长期进化中除了产生逃逸或抑制天然免疫应答的能力外,还拥有多种不同机制逃逸或抑制获得性免疫应答。病原突破机体天然免疫应答屏障建立感染后,机体主要通过获得性免疫来抑制和清除病原。机体获得性免疫应答主要通过抗体介导(特别是中和抗体)的体液免疫和细胞毒性T 细胞(cytotoxic T-lymphocyte,CTL)介导的细胞免疫来发挥作用。然而,病原可通过遗传变异和重组产生新的变异毒株,从而逃逸获得性免疫应答中的中和抗体和 CTL。病毒基因组,特别是 RNA 病毒,在免疫选择压力下可不断地快速突变以适应宿主系统。例如流感病毒和冠状病毒可通过抗原漂移来逃避 T 细胞和病毒中和抗体的识别,削弱机体获得性免疫保护性,其中一些变异毒株还可获得比原始毒株更强大的传播能力或毒力,如当前新冠病毒的奥密克戎(Omicron)变异株的传播能力较武汉原始毒株显著增加,Omicron BA.4/5 R0 从武汉原始毒株的 3.3 增加到 18.6,并且由于 spike 蛋白突变导致现有疫苗的保护力大大降低,这也是导致疫情反复流行的重要原因之一。
除了被动逃逸机体免疫应答,一些重要病原能够通过多种机制主动攻击宿主免疫系统从而抑制宿主免疫应答,促进感染。很多病原能够感染机体重要免疫器官,如猪繁殖与呼吸综合征病毒(PRRSV) 和猪圆环病毒能够感染胸腺与骨髓,机体分别负责 T 细胞和 B 细胞发育的重要免疫器官,导致免疫器官功能损伤,抑制机体获得性免疫应答产生,从而建立持续感染。此外,PRRSV 和猪伪狂犬病毒还可通过干扰抗原加工和呈递达到抑制诱导获得性免疫应答。
病原体和宿主的长期博弈就像一场无休止的进化军备竞赛。人类要想在这场军备竞赛中取得胜利,必须知己知彼,继续加大对相关领域的研究和投入,突破传统模式,从病原和宿主两方面进行创新性构想。① 不同物种免疫系统差异非常大,需继续加大对畜禽免疫学基础研究,揭示机体诱导免疫应答的一些重要分子和信号通路,以及重要病原感染后免疫保护的分子机制。② 通过基因编辑技术构建基因缺失毒株,进一步揭示病原介导免疫逃逸或免疫抑制的关键基因 , 在此基础上构建弱毒疫苗株。③ 研发广谱性疫苗。利用不断发展的新技术研发广谱性疫苗是有效防控当前高度变异重要病原最有效、最经济的方法。尽管病毒比宿主进化得更快,但病毒的重要蛋白质受到许多影响病毒复制的功能限制,存在一些保守位点。大量研究已经证实易发生变异的病毒都存在广谱性中和抗体和T细胞表位。利用单个细胞测序技术和高通量筛选技术鉴定出广谱性中和抗体和 CD8+T 细胞识别位点, 以及利用生物信息学和核酸合成技术设计合成保守抗原基因的广谱性 mRNA 或 DNA 疫苗,可为未来开发广谱性疫苗提供重要指导。
在“重要动物病原的免疫抑制与逃逸机制”工程研究前沿中,核心论文数排名前三位的国家分别是美国(占 51.11%)、英国(占 31.11%)和中国(占17.78%)(表 1.2.5)。该前沿的核心论文被引频次分布在 240~3 616,篇均被引频次均超过 100, 其中英国篇均被引频次超过 200,泰国和丹麦的篇被引频次均超过了 300。研究机构分布方面,牛津大学、南安普敦大学和剑桥大学产出的核心论文及被引次数较多(表 1.2.6)。主要国家间的合作网络(图 1.2.4)方面,国家间的研究合作较为普遍, 以英国和美国合作相对更紧密。主要机构间的合作网络(图 1.2.5)方面,各机构间均存在一定的合作关系。施引核心论文的主要产出国家主要是美国和中国,美国占比接近 1/3, 中国超过 10%,且平均施引年较晚,表现出较强的研发后劲(表 1.2.7)。施引核心论文的主要产出机构为中国科学院、哈佛
《表 1.2.5》
表 1.2.5 “重要动物病原的免疫抑制与逃逸机制”工程研究前沿中核心论文的主要产出国家
| 序号 | 国家 | 核心论文数 | 论文比例 /% | 被引频次 | 篇均被引频次 | 平均出版年 |
| 1 | 美国 | 23 | 51.11 | 3 063 | 133.17 | 2020 |
| 2 | 英国 | 14 | 31.11 | 3 616 | 258.29 | 2020.4 |
| 3 | 中国 | 8 | 17.78 | 1 156 | 144.5 | 2019.9 |
| 4 | 瑞士 | 4 | 8.89 | 554 | 138.5 | 2021 |
| 5 | 荷兰 | 4 | 8.89 | 510 | 127.5 | 2019.5 |
| 6 | 南非 | 3 | 6.67 | 438 | 146 | 2021 |
| 7 | 泰国 | 2 | 4.44 | 774 | 387 | 2020.5 |
| 8 | 丹麦 | 2 | 4.44 | 681 | 340.5 | 2021 |
| 9 | 印度 | 2 | 4.44 | 300 | 150 | 2021 |
| 10 | 新加坡 | 2 | 4.44 | 240 | 120 | 2020.5 |
《表 1.2.6》
表 1.2.6 “重要动物病原的免疫抑制与逃逸机制”工程研究前沿中核心论文的主要产出机构
| 序号 | 机构 | 核心论文数 | 论文比例 /% | 被引频次 | 篇均被引频次 | 平均出版年 |
| 1 | 牛津大学 | 8 | 17.78 | 2 202 | 275.25 | 2020.5 |
| 2 | 南安普敦大学 | 5 | 11.11 | 1 493 | 298.6 | 2020.2 |
| 3 | 剑桥大学 | 4 | 8.89 | 1 183 | 295.75 | 2020.2 |
| 4 | 得克萨斯大学奥斯汀分校 | 3 | 6.67 | 879 | 293 | 2020.3 |
| 5 | 哈佛大学 | 3 | 6.67 | 570 | 190 | 2021 |
| 6 | 爱丁堡大学 | 2 | 4.44 | 1 101 | 550.5 | 2021 |
| 7 | 格拉斯哥大学 | 2 | 4.44 | 775 | 387.5 | 2021 |
| 8 | 维康桑格研究所 | 2 | 4.44 | 739 | 369.5 | 2021 |
| 9 | 帝国理工学院 | 2 | 4.44 | 681 | 340.5 | 2021 |
| 10 | 哥本哈根大学 | 2 | 4.44 | 681 | 340.5 | 2021 |
《图 1.2.4》
图 1.2.4 “重要动物病原的免疫抑制与逃逸机制”工程研究前沿主要国家间的合作网络
《图 1.2.5》
图 1.2.5 “重要动物病原的免疫抑制与逃逸机制”工程研究前沿主要机构间的合作网络
《表 1.2.7》
表 1.2.7 “重要动物病原的免疫抑制与逃逸机制”工程研究前沿中施引核心论文的主要产出国家
| 序号 | 国家 | 施引核心论文数 | 施引核心论文比例 /% | 平均施引年 |
| 1 | 美国 | 1 835 | 31.7 | 2020.7 |
| 2 | 中国 | 1 050 | 18.14 | 2020.7 |
| 3 | 英国 | 555 | 9.59 | 2020.7 |
| 4 | 印度 | 419 | 7.24 | 2020.7 |
| 5 | 德国 | 404 | 6.98 | 2020.8 |
| 6 | 意大利 | 354 | 6.12 | 2020.7 |
| 7 | 法国 | 283 | 4.89 | 2020.7 |
| 8 | 巴西 | 263 | 4.54 | 2020.8 |
| 9 | 澳大利亚 | 235 | 4.06 | 2020.7 |
| 10 | 加拿大 | 218 | 3.77 | 2020.7 |
大学和牛津大学,中国科学院的施引论文量排在首位(表 1.2.8)。图 1.2.6 为“重要动物病原的免疫抑制与逃逸机制”工程研究前沿的发展路线。
1.2.3 土壤高效固碳与调控机制
土壤高效固碳与调控研究的重要意义。土壤有机质是土壤健康的核心,维系着土壤肥力,是保障粮食安全的基础,且与生态系统生产力和功能可持续性息息相关。此外,土壤作为重要的碳封存场所,土壤有机碳库是陆地生态系统中最大的碳库,全球土壤有机碳总量为 1 500~2 000 Pg,相当于大气中碳总量的 2~3 倍。研究发现农业土壤碳固定占自然气候解决方案总潜力的 25%。因此,促进农业土壤固碳对于保障粮食安全、减缓气候变化以及推进农业绿色发展具有重要意义。土壤高效固碳与调控的核心是深入认识土壤有机质的形成、周转和稳定等关键过程,建立高效固碳、减缓矿化分解的调控机制。
土壤高效固碳与调控机制研究现状。土壤有机质的形成和稳定过程具有高度复杂性。目前对土壤有机质的形成过程、赋存形态和稳定机理等认知仍然不足并存在分歧。经典腐殖化理论所定义的腐殖
《表 1.2.8》
表 1.2.8 “重要动物病原的免疫抑制与逃逸机制”工程研究前沿中施引核心论文的主要产出机构
| 序号 | 机构 | 施引核心论文数 | 施引核心论文比例 /% | 平均施引年 |
| 1 | 中国科学院 | 263 | 22.87 | 2020.8 |
| 2 | 哈佛大学 | 147 | 12.78 | 2020.8 |
| 3 | 牛津大学 | 146 | 12.7 | 2020.7 |
| 4 | 华盛顿大学 | 100 | 8.7 | 2020.7 |
| 5 | 剑桥大学 | 77 | 6.7 | 2020.8 |
| 6 | 香港大学 | 73 | 6.35 | 2020.6 |
| 7 | 复旦大学 | 73 | 6.35 | 2020.7 |
| 8 | 帝国理工学院 | 70 | 6.09 | 2020.8 |
| 9 | 圣保罗大学 | 68 | 5.91 | 2020.8 |
| 10 | 西奈山伊坎医学院 | 68 | 5.91 | 2020.7 |
《图 1.2.6》
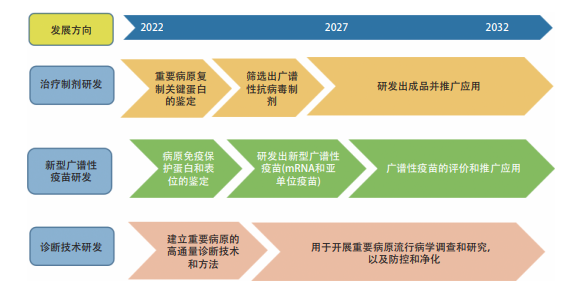
图 1.2.6 “重要动物病原的免疫抑制与逃逸机制”工程研究前沿的发展路线
质具有高度复杂性和模糊性,传统的研究手段尚不足以建立较明确的“白箱”模型。目前关于有机质形成和稳定的最新学说是有机质连续体模型,即植物残体向土壤有机质的转化是从大的植物生物聚合物到小分子化合物的微生物逐级分解过程,因而土壤有机质的存在形式是从大的植物碎片到逐渐分解成的小分子化合物的连续体。在该模型中,外源有机物料在被微生物利用的过程中体积不断减小,热动力学梯度逐渐下降,而极性组分、可溶性组分和离子化组分相应增加。并且,随着分子复杂程度的逐渐下降,有机化合物更易于与矿物表面结合或进入团聚体内部而增加其稳定性。但也有研究认为尽管土壤微生物可将腐殖质完全或部分分解,但同时会产生新的腐殖质使有机质得以更新。总之,传统的腐殖化理论和有机质连续体模型均承认动植物碎片在输入土壤后,会先经过物理化学作用而破碎, 进而通过胞外酶等降解成相对更小的组分,最终在土壤中固存。
有机质形成和稳定过程与周围土壤基质相互作用密切相关。土壤团聚体是土壤有机质主要的赋存场所,能通过自身的物理保护作用将有机质包被起来,从而免受微生物的分解。因此,团聚体保护能力和容量是土壤固碳潜力的物理基础。土壤对有机质的化学保护作用主要指土壤无机分子与有机分子之间的相互作用而使有机质难以被微生物利用。新的研究发现,微生物可通过同化作用将土壤中可利用碳源以代谢产物的形式贡献于土壤有机质,在农业土壤中,其贡献可占有机碳的 50% 以上。此外,其还能通过影响团聚体的周转而间接作用于土壤有机质稳定性。因此,目前研究认为,有机质的稳定性是团聚体的物理保护– 土壤矿物的结合 – 微生物代谢过程相互作用、相互依存的结果。
合理的管理措施可以通过影响碳输入与输出之间的平衡,进而调控土壤固碳。外源碳输入是土壤有机质形成的重要来源,土壤中微生物的代谢活动可以将植物残体转化为土壤有机质。传统耕作模式有机物料补给不足、土壤翻动频繁,导致土壤团聚体结构被破坏、有机碳损耗加快,从而限制微生物的生长和代谢,不利于土壤有机质的累积。此外, 多样化种植及轮作改变了作物残体的数量和质量, 显著影响土壤微生物群落结构和活性,进而提高了土壤有机质的含量。
未来研究方向和创新点。土壤是一个多种物质并存、多种过程同时发生、多种因素共同影响的开放的、复杂的系统。目前土壤高效固碳机理还存在很多不清楚的地方,土壤有机质提升困难。因此未来需要强化对固碳过程和调控机制的深入研究。如:精确区分不同有机组分碳源、探究植物残体向土壤有机质转化的微生物作用机制、加快基于多因素协同调控的有机质稳定性研究、探索土壤碳平衡机理及植物残体输入阈值等,强化对土壤固碳本质和调控机制的研究。创新高效固碳保肥新型碳基材料,提高土壤固碳效率、集成创新土壤固碳、耕地保育及产能提升关键技术及技术模式,推动大面积农业土壤固碳落地应用为该领域的重要应用研究前沿(图 1.2.7)。
《图 1.2.7》
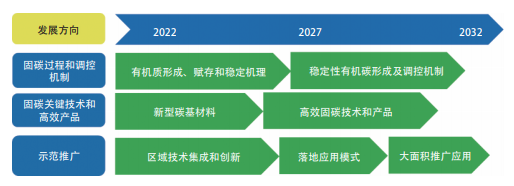
图 1.2.7 “土壤高效固碳与调控机制”工程研究前沿的发展路线
在“土壤高效固碳与调控机制”工程研究前沿中,核心论文数排在前三位的国家分别是中国、美国和德国(表 1.2.9)。该前沿的核心论文篇均被引频次分布在 94.80~297.75,除沙特阿拉伯外,其他国家篇被引频次均超过了 100。研究机构分布方面(表 1.2.10),兰州大学、苏黎世联邦理工学院、苏塞克斯大学、清华大学产出的核心论文及被引次数较多。施引核心论文的主要产出国家是中国、美国和澳大利亚(表 1.2.11)。在施引核心论文的主要产出机构(表 1.2.12)方面,中国科学院大学、湖南大学、清华大学施引论文数排在前三位。国家间的合作较为普遍、网络复杂(图 1.2.8),中国、美国、德国和英国合作相对更为紧密。主要机构间的合作网络(图 1.2.9)方面,同一国家内的不同机构及不同国家的各机构间均存在一定的合作关系,如兰州大学与中国科学院,清华大学与韩国大学,苏黎世联邦理工学院、萨塞克斯大学和莫纳什大学间均有紧密合作。
《表 1.2.9》
表 1.2.9 “土壤高效固碳与调控机制” 工程研究前沿中核心论文的主要产出国家
| 序号 | 国家 | 核心论文数 | 论文比例 /% | 被引频次 | 篇均被引频次 | 平均出版年 |
| 1 | 中国 | 51 | 61.45 | 9 480 | 185.88 | 2019.1 |
| 2 | 美国 | 32 | 38.55 | 5 545 | 173.28 | 2018.5 |
| 3 | 德国 | 15 | 18.07 | 2 954 | 196.93 | 2018.3 |
| 4 | 英国 | 13 | 15.66 | 2 702 | 207.85 | 2018.5 |
| 5 | 澳大利亚 | 10 | 12.05 | 1 821 | 182.1 | 2018.5 |
| 6 | 韩国 | 9 | 10.84 | 1 267 | 140.78 | 2018.6 |
| 7 | 奥利地 | 5 | 6.02 | 1 061 | 212.2 | 2018 |
| 8 | 沙特阿拉伯 | 5 | 6.02 | 474 | 94.8 | 2019.6 |
| 9 | 瑞士 | 4 | 4.82 | 1 191 | 297.75 | 2018.5 |
| 10 | 法国 | 4 | 4.82 | 1 098 | 274.5 | 2019.5 |
《表 1.2.10》
表 1.2.10 “土壤高效固碳与调控机制” 工程研究前沿中核心论文的主要产出机构
| 序号 | 机构 | 核心论文数 | 论文比例 /% | 被引频次 | 篇均被引频次 | 平均出版年 |
| 1 | 湖南大学 | 5 | 6.02 | 782 | 156.4 | 2018 |
| 2 | 韩国大学 | 4 | 4.82 | 605 | 151.25 | 2018.5 |
| 3 | 中国科学院 | 4 | 4.82 | 480 | 120 | 2019.5 |
| 4 | 沙特国王大学 | 4 | 4.82 | 257 | 64.25 | 2019.5 |
| 5 | 兰州大学 | 3 | 3.61 | 1 227 | 409 | 2018.3 |
| 6 | 苏黎世联邦理工学院 | 3 | 3.61 | 1 079 | 359.67 | 2018.3 |
| 7 | 萨塞克斯大学 | 3 | 3.61 | 971 | 323.67 | 2019 |
| 8 | 清华大学 | 3 | 3.61 | 852 | 284 | 2018.7 |
| 9 | 马萨诸塞大学 | 3 | 3.61 | 784 | 261.33 | 2016.7 |
| 10 | 莫纳什大学 | 3 | 3.61 | 763 | 254.33 | 2019.7 |
《表 1.2.11》
表 1.2.11 “土壤高效固碳与调控机制” 工程研究前沿中施引核心论文的主要产出国家
| 序号 | 国家 | 施引核心论文数 | 施引核心论文比例 /% | 平均施引年 |
| 1 | 中国 | 1 492 | 43.9 | 2019.3 |
| 2 | 美国 | 522 | 15.36 | 2019.1 |
| 3 | 澳大利亚 | 240 | 7.06 | 2019.3 |
| 4 | 英国 | 236 | 6.94 | 2019.4 |
| 5 | 德国 | 225 | 6.62 | 2019.1 |
| 6 | 韩国 | 148 | 4.35 | 2019.4 |
| 7 | 加拿大 | 123 | 3.62 | 2019.4 |
| 8 | 印度 | 116 | 3.41 | 2019.8 |
| 9 | 西班牙 | 115 | 3.38 | 2019.1 |
| 10 | 荷兰 | 96 | 2.82 | 2019.3 |
《表 1.2.12》
表 1.2.12 “土壤高效固碳与调控机制” 工程研究前沿中施引核心论文的主要产出机构
| 序号 | 机构 | 施引核心论文数 | 施引核心论文比例 /% | 平均施引年 |
| 1 | 中国科学院大学 | 246 | 26.59 | 2019 |
| 2 | 湖南大学 | 160 | 17.3 | 2019 |
| 3 | 清华大学 | 123 | 13.3 | 2019.3 |
| 4 | 韩国大学 | 62 | 6.7 | 2019.5 |
| 5 | 香港理工大学 | 58 | 6.27 | 2019.5 |
| 6 | 兰州大学 | 48 | 5.19 | 2017.9 |
| 7 | 苏黎世联邦理工学院 | 47 | 5.08 | 2018.9 |
| 8 | 韩国世宗大学 | 47 | 5.08 | 2019.4 |
| 9 | 浙江大学 | 47 | 5.08 | 2019.1 |
| 10 | 哈尔滨工业大学 | 44 | 4.76 | 2019.2 |
《图 1.2.8》
图 1.2.8 “土壤高效固碳与调控机制”工程研究前沿主要国家间的合作网络
《图 1.2.9》
图 1.2.9 “土壤高效固碳与调控机制”工程研究前沿主要机构间的合作网络
《2 工程开发前沿》
2 工程开发前沿
《2.1 Top 11 工程开发前沿发展态势》
2.1 Top 11 工程开发前沿发展态势
农业领域的 Top 11 工程开发前沿主要涉及生物育种、智慧农业、绿色农业等方向(表 2.1.1)。其中,与生物育种相关的开发前沿包括动物精准基因编辑育种技术、智能制种技术、园艺作物基因编辑技术应用、林木全基因组选择育种、基于 RNA 干扰的病虫害防控技术;与智慧农业相关的开发前沿包括农业自主作业机器人、作物无人化智慧栽培技术、生态智能池塘养殖技术;与绿色农业相关的开发前沿包括有机污染物催化降解技术、饲用抗生素替代技术与产品;同时,新发和再现重大动物疫病监测与预警也是科研人员热衷的研究对象。其中,智能制种技术的平均被引数高达67.69,说明智能制种技术近年来得到了科研人员的广泛关注。
各前沿涉及的核心专利 2016—2020 年公开情况见表 2.1.2,其中动物精准基因编辑育种技术的核心专利最多,2021 年更是高达 829 项,远高于其他开发前沿。林木全基因组选择育种的核心专利最少,只有 2017 年、2021 年出现过核心专利。
(1)新发和再现重大动物疫病监测与预警
“新发和再现重大动物疫病”是指新出现或再度肆虐的且造成危害巨大的动物传染病,其包括原有病原体变异或演化导致的传染病、原先未被认识的传染病、扩散到新的地理区域或动物群体的已知传染病、已被控制但因抗药性改变或防控措施削弱而重新出现或再度流行的传染病。近年来境外新传入动物疫病达 30 余种,非洲猪瘟、小反刍兽疫和牛结节性皮肤病等重大疫病相继传入,特别是非洲猪瘟席卷全国,重创我国养猪业。同时,境外动物疫情呈多发态势,非洲马瘟、疯牛病等在“一带一路”沿线国家和地区扩散与蔓延,跨境传入风险巨大。该类重大动物疫病不仅严重威胁畜禽养殖生产安全、经济安全和生态安全,部分疫病还具有人兽共患风险。联合国粮食及农业组织(FAO)指出, “强大的国际和国家动物卫生体系是预防疫病、确保安全营养食物、保护农民利益的关键”。因此, 新发和再现重大动物疫病监测与预警对于保障国家生物安全,预防与控制疫病暴发与流行至关重要。其核心技术需求主要包括:① 快速、准确、便捷、高通量、可鉴别诊断的检测技术创新与集成;② 明确疫病病原学基本特性、传播规律、分布特征、分子演化路径、与宿主协同进化关系以及暴发机理,
《表 2.1.1》
表 2.1.1 农业领域 Top 11 工程开发前沿
| 序号 | 工程开发前沿 | 公开量 | 引用量 | 平均被引数 | 平均公开年 | |
| 1 | 新发和再现重大动物疫病监测与预警 | 454 | 1 186 | 2.61 | 2018.7 | |
| 2 | 动物精准基因编辑育种技术 | 4 546 | 10 294 | 2.26 | 2018.2 | |
| 3 | 智能制种技术 | 88 | 5 957 | 67.69 | 2016.7 | |
| 4 | 园艺作物基因编辑技术应用 | 50 | 121 | 2.42 | 2019.3 | |
| 5 | 林木全基因组选择育种 | 2 | 7 | 3.5 | 2019 | |
| 6 | 基于 RNA 干扰的病虫害防控技术 | 94 | 913 | 9.71 | 2018.4 | |
| 7 | 农业自主作业机器人 | 259 | 861 | 3.32 | 2019.3 | |
| 8 | 有机污染物催化降解技术 | 1 000 | 2 039 | 2.04 | 2018.4 | |
| 9 | 饲用抗生素替代技术与产品 | 108 | 140 | 1.3 | 2018.4 | |
| 10 | 作物无人化智慧栽培技术 | 261 | 720 | 2.76 | 2018.9 | |
| 11 | 生态智能池塘养殖技术 | 103 | 182 | 1.77 | 2019 | |
《表 2.1.2》
表 2.1.2 农业领域 Top 11 工程开发前沿核心专利逐年公开量
| 序号 | 工程开发前沿 | 2016 | 2017 | 2018 | 2019 | 2020 | 2021 |
| 1 | 新发和再现重大动物疫病监测与预警 | 78 | 62 | 62 | 76 | 68 | 108 |
| 2 | 动物精准基因编辑育种技术 | 433 | 654 | 697 | 814 | 685 | 829 |
| 3 | 智能制种技术 | 4 | 9 | 10 | 14 | 9 | 15 |
| 4 | 基因编辑技术应用 | 1 | 4 | 11 | 9 | 13 | 12 |
| 5 | 林木全基因组选择育种 | 0 | 1 | 0 | 0 | 0 | 1 |
| 6 | 基于 RNA 干扰的病虫害防控技术 | 10 | 20 | 19 | 22 | 12 | 11 |
| 7 | 农业自主作业机器人 | 9 | 28 | 48 | 43 | 56 | 63 |
| 8 | 有机污染物催化降解技术 | 68 | 142 | 240 | 191 | 211 | 95 |
| 9 | 饲用抗生素替代技术与产品 | 16 | 10 | 13 | 11 | 22 | 24 |
| 10 | 作物无人化智慧栽培技术 | 22 | 28 | 36 | 47 | 48 | 72 |
| 11 | 生态智能池塘养殖技术 | 3 | 16 | 25 | 8 | 13 | 35 |
依据病原分子生态学建立疾病发生和扩散模型,提出相应的疾病暴发预测指标;③ 建立并完善动物重大疫病信息数据库、预警与报告系统,提升对发病概率和潜在危害的精准评估能力。相关技术突破将为国家重大动物疫病的防控和生物安全预警提供技术支撑和解决方案。
(2)动物精准基因编辑育种技术
动物育种技术经历了从最初的表型值育种技术到现代的基因组编辑育种技术的跨越式发展。传统的杂交改良方法成本高、耗时长、过程复杂,还可能因基因连锁引入缺陷基因,而基因编辑技术则可以对目的基因进行精确修饰,通过修饰动物重要性状相关的关键功能基因和调控序列,一代即可获得具有特性表型的动物新品种(系),从而提高动物育种效率。基因编辑技术自问世以来已经发展更新 3 代:锌指核酸酶(ZFNs)编辑技术、类转录激活因子效应物核酸酶(TALENs)编辑技术和CRISPR-Cas9 编辑技术。基因编辑技术由最先对目标基因的随机打靶失活到现在可以实现对基因单个碱基的精准替换,技术上已经实现了重大突破。近几年来,基因编辑技术已成为动物育种的重要方法, 在动物育种领域得到十足的发展和应用。已通过基因编辑技术精准创制了一批高产、优质、高繁殖和抗病育种新材料和新品系,如抗繁殖与呼吸综合征的基因编辑猪、肌肉生长抑素(MSTN)基因编辑的高产肉牛和抗结核病奶牛等。随着多组学测序技术的不断进步,各种表型和基因型数据库不断丰富, 多种性状相关的重要功能基因和调控序列挖掘不断深入,基因编辑技术的效率、准确性和安全性不断提升,动物精准基因编辑育种技术将逐渐向工程化和规模化方向发展,未来将在动物育种领域广泛应用,为动物新品种培育提供重要技术支撑。
(3) 智能制种技术
自 20 世纪中期开始,玉米水稻等作物杂交种在生产中的普及使用是杂种优势提升农业生产水平的一大革命性标志。杂交种的生产可以通过利用母本雄性不育系或对母本人工去雄来实现。然而,在生产杂交种的过程中,由于环境的变化可能导致雄性不育或去雄不彻底,从而导致母本自交授粉,致使生产的杂交种中混杂了母本自交系的种子,母本自交系的产量远低于杂交种的产量,这为种子生产带来了重要的潜在威胁。同时,传统不育制种技术依赖长周期的导育和合适的配套恢复系,去雄则附加了大量的人力和时间成本。建立新一代杂交种智能制种技术,一方面能够大大节约制种成本,另一方面还能够潜在地提升杂交种的群体产量。目前智能制种研究领域经多年技术优化,已经初步建立了基于种子大小、荧光、种皮及植物色素等筛选标记的大规模、高精度核不育系的技术体系,在此基础上,结合多个不育基因的利用,继续优化和改进该技术,将为解决我国杂交种制种技术更新换代提供新的解决方案。
(4) 园艺作物基因编辑技术应用
基因编辑是通过敲除几个碱基对或一段 DNA序列从而改变原有基因序列的技术,主要利用的 3 种工具酶分别为 ZFNs、TALENs 和 CRISPR- Cas9,其中近年诞生的 CRISPR-Cas9 基因编辑系统由于编辑效率高、操作简单、成本低等优点,近年来成为基因编辑炙手可热的工具,在植物中均得到广泛应用。基因编辑技术由最先对目标基因的随机打靶失活到现在可以实现对基因单个碱基的精准替换,技术上已经实现了重大突破,为基因功能研究及作物性状改良做出了重要贡献。2021 年,一种经过 CRISPR 技术改造的富含 γ- 氨基丁酸的番茄在日本上市销售。基因编辑技术在作物和模式植物中已得到广泛应用,基因编辑技术在作物遗传改良上相较于传统杂交育种有很大的优势。而园艺作物种类繁多,目前已在番茄等园艺作物建立了较为成熟的基因编辑体系,但很多的园艺作物尚未有较好的基因编辑体系,在不同的园艺作物中建立高效的基因编辑体系还需要大量的实践探索。
(5) 林木全基因组选择育种
林木全基因组选择育种属于林业科学学科, 是林木育种的新兴开发前沿技术。林木重要性状多为复杂的数量性状、遗传杂合性高,常规育种技术难以高效快速定向培育林木良种。分子标记辅助选择育种(molecular marker assistant selection breeding,MAS)难以捕获微效位点、遗传作图分辨率低,数量性状遗传定位结果间难以互相验证, 不能有效推动林木遗传改良进程。全基因组选择(genomic selection,GS)是指利用覆盖全基因组的高密度遗传标记对复杂数量性状进行预测的育种方法,能够克服 MAS 利用少量标记选择育种的不足,极大地提高对微效位点的捕获功效。主要技术环节包括:① 建立训练群体,测定所有个体的表型和基因型,利用合适的统计模型估计单核苷酸多态性位点效应值,建立表型和基因型间的 GS 预测模型;② 基于候选群体中个体基因型数据,利用GS 预测模型计算候选群体个体基因组估计育种值(genome estimated breeding value,GEBV);③ 根据 GEBV 排序在苗期筛选个体,测定表型后再纳入训练群体持续更新和优化 GS 模型,增强预测精度和功效;④ 促进林木 GS 与新兴技术(如早花、体细胞胚发生、基因编辑等)融合,真正实现利用基因组信息指导育种实践。林木全基因组选择育种可在林木幼苗甚至种子阶段开展优良基因型选育, 增强遗传增益、加快选育进程、促进林木精准高效育种,是具有巨大潜力的林木育种策略。
(6) 基于 RNA 干扰的病虫害防控技术
RNA 干扰现象是一项获得诺贝尔生理学或医学奖(2006 年)、具有跨时代伟大意义的发现。RNA 干扰技术已经在医药、农业等多个领域展现出巨大的应用前景,截至 2022 年,全球已有近 20 种基于 RNA 干扰的医药新产品进入临床研究阶段。在农业领域,RNA 干扰可以沉默有害生物生长发育或重要过程中的关键基因,阻碍有害生物正常的生长和繁殖,导致其死亡,从而降低有害生物的危害程度。基于 RNA 干扰的病虫害防控技术被称为“农药史上的第三次革命”,该技术具有靶标专一性强、无毒、无残留等特点,在发展环境友好型农业中具有跨时代意义。2017 年,美国环境保护署批准了全球第一款基于 RNA 干扰的抗虫转基因玉米——MON87411(拜耳公司),该产品于2020 年获得中国农业农村部转基因安全许可证书。此外,已有多种 RNA 生物农药已提交 EPA 审核, 如 BioDirect(拜耳公司)、Ledprona GS2(绿光生物科学)等。我国关于 RNA 干扰的病虫害防控技术研发起步较早,但商品化应用进程缓慢,相关的技术标准、法律法规也不完善。此外,在 RNA 干扰产品的商品化研发过程中,有几个关键的核心问题亟待解决:① RNA 干扰抗病虫靶标基因的筛选;② 双链 RNA 工业化合成方法的开发;③ 双链RNA 递送或缓释载体的构建。尽快解决上述问题, 可以推进我国新型RNA 抗病虫技术商品化的进程, 有助于我国追赶国际前沿科技、抢占国际市场、树立我国在该领域的国际地位。
(7) 农业自主作业机器人
农业自主作业机器人是具有感知、决策、控制和执行的智能农机装备。农业自主作业机器人是机器人领域的重要分支,是农业机械、人工智能、机器人和信息工程等技术交叉融合领域,是现代农业发展的重要趋势。当前,我国农业综合机械化率已超过 70%,农业机械化解放了劳动力、提高了劳动生产率和资源利用率、促进了农民增收。然而, 随着我国城镇化进程加快和农村劳动力大量进城务工,农村劳动力出现了季节性和结构性短缺,农业自主作业机器人将是解决“谁来种地”和“怎样种地” 的重要农机装备。针对非结构化复杂农业场景,农业自主作业机器人重点突破如下关键理论与技术:① 作业机器人、作业环境和作业对象的信息精准感知机理和传感技术;② 基于多源感知异构信息的物景认知方法,以及基于作业流程和机器学习的自主决策、规划与控制技术;③ 融合先进农艺的作业装置、末端执行器和机械臂先进设计与精准高效作业技术;④ 全地形、多遮挡和动态农业场景下的底盘结构设计与线控技术,以及多传感融合的地图构建、自主规划与避障导航技术;⑤ 机器人群体实时通信、群体自主协同和人机共融技术。农业自主作业机器人代表着最先进的农业生产力,将最大程度地解放农业劳动力,极大地提高生产率、资源利用率和产出率,实现农业智慧生产。
(8) 有机污染物催化降解技术
有机污染物种类繁多、成分复杂、结构多样, 具有环境迁移性、生物积累性和生态毒性,成为农业环境领域的新兴关注点。有机污染物的赋存影响农田土壤环境质量,并给城乡生活废水处理及农业农村有机废弃物资源化利用带来挑战。开发经济、高效的有机污染物催化降解技术,是当前的研究热点和应用前沿。现阶段备受关注的有机污染物催化降解技术主要包括光催化降解技术、非均相催化过硫酸盐降解技术、臭氧催化氧化技术、电催化降解技术等。尽管催化降解技术去除有机污染物效率较高,但在实际应用中易受共存物质的干扰,创新特异性目标有机污染物专项去除的催化降解技术是该方向的重要研究趋势之一。此外,有机物催化降解的中间产物会影响催化降解效率,且可能对生态系统构成威胁,因此定向调控有机污染物催化降解的中间产物是该方向的另一重要需求。因此,提高有机污染物的矿化效率,揭示其催化降解途径及降解机理,开发节能高效的催化降解技术及设备,是该领域亟须突破的重点和难点。
(9) 饲用抗生素替代技术与产品
抗生素的大量、长期使用不仅造成了严重的生态环境污染,更使一些病原微生物产生了耐药性, 造成“超级细菌”的出现,危及人类和动物的健康。我国已于 2020 年 7 月 1 日起正式禁止饲用抗生素的使用。从欧洲的“禁抗”历程来看,饲料全面“禁抗” 后养殖业会出现畜禽发病率、死亡率提高,养殖成本增加等问题。开发高效、环保、无毒副作用的饲用抗生素替代品,提高畜禽自身免疫抗病力,有利于促进我国养殖业健康发展。研究较多的饲用抗生素替代品有酸化剂、抗菌肽、植物精油等。益生菌、益生元、抗菌肽、植物提取物、中草药以及酶制剂等已被证明具有较好的调控畜禽肠道微生态以及改善肠道健康的效果,在无抗养殖发展过程中具有广泛的前景。合成生物学在这些产品的创制过程中发挥了重要作用。目前,以合成生物学为核心的第三代生物技术已经进入一个技术日新月异、产业蓬勃发展的新阶段。经济合作与发展组织(OECD)预测, 至 2030 年,35% 的化学品和其他工业产品将来自生物制造,奠定了生物制造产业作为全球战略新兴产业的核心位置。以合成生物学为指导,发展基于微生物的农业生物制造技术,设计饲用生物活性分子的高效合成路线和人工生物体,突破自然生物体合成的局限,重塑生物饲料生产方式,创制新型饲料资源和饲料添加剂并实现产业化,可有效解决我国饲料原料短缺、抗生素超量添加及畜牧产品安全的问题,将产生广泛的社会效益和经济效益。
(10) 作物无人化智慧栽培技术
现代科技的迅猛发展以及农业生产经营方式的升级转型使无人化智慧栽培成为作物生产未来发展的基本方向。世界各国依靠经济与科技的投入正向作物无人化智慧栽培方向进军。作物无人化智慧栽培,即以绿色优质丰产高效生产为目标,利用卫星导航系统、物联网、大数据、人工智能等新一代信息技术与现代作物栽培技术相融合,通过智能机具(设备)代替人实施作物田间耕种收精确化作业、作物生长精准化监测诊断以及水肥药精准变量施用,实现全程栽培作业无人化与作物稳健生育,具有精准智能、自动高效、安全可靠、多能通用等特点, 是作物生产方式的重大颠覆性变革。作物无人化智慧栽培是一项作物栽培、机械工程、信息科学等多领域深度融合的系统工程技术,其核心科学问题主要有:① 作物无人化智慧栽培并能实现绿色优质丰产高效协同生产的有效途径与机制;② 适合无人化智慧化栽培系统的种、肥、药等产品的研制与筛选;③ 应用于无人化智慧栽培的融合信息技术的农机装备与设施设备的研制及选型配套;④ 作物绿色优质丰产高效的“栽培 – 机械 – 信息”深度融合的栽培技术开发与应用。例如,以稻麦轮作为对象,通过多学科协同创新,创建绿色丰产优质高效协同的关键栽培农艺与全程化最轻简无人化智慧栽培作业模式,周年生产无人化智慧化率达 85%以上,周年均产稳定达每亩 1 1 300 kg 以上,实现大规模推广应用,推动稻麦轮作无人化智慧栽培达到国际领跑水平。
(11) 生态智能池塘养殖技术
目前,我国水产养殖面积约为 70 360 km2,其中淡水养殖面积为 50 410 km2(占 71.6%),池塘养殖是淡水养殖的主要方式(占 74%),池塘养殖业在淡水养殖业甚至是全球水产养殖业中都占有举足轻重的地位,是稳定水产品供给的基础。池塘养殖的健康可持续发展涉及国家粮食安全,亟须持续且稳步地推进发展。池塘养殖面临的关键问题包括种源缺乏、池塘老旧、配套技术落后等。针对这些问题,国内外学者通过制备优质养殖苗种、研发池塘养殖水质精准调控技术、开发高效净化装备与减排系统、建立以物联网为基础的全程信息化养殖管控系统,提升传统池塘养殖智能及精准控制水平, 最终建立生态化、智能化、工业化的现代池塘养殖模式,推动了现代池塘养殖的健康可持续发展。
1 1 亩≈ 667 m2。
《2.2 Top 3 工程开发前沿重点解读》
2.2 Top 3 工程开发前沿重点解读
2.2.1 动物精准基因编辑育种技术
动物精准基因编辑育种技术是指利用 CRISPR- Cas9 等基因组编辑工具,精确修饰动物重要性状相关功能基因和调控序列,定向培育具有特定表型的动物新品种(系)的方法。基因编辑技术自问世以来已经发展更新 3 代:ZFNs 编辑技术、TALENs 编辑技术和 CRISPR-Cas9 编辑技术。由最先对目标基因的剪切断裂引入突变,到现在可以实现对基因单个碱基的精准替换,以及大片段的精确插入或者替换等操作,技术上已经实现了重大突破。随着多组学测序技术的不断进步,各种表型和基因型数据库不断丰富,多种性状相关的重要功能基因和调控序列挖掘不断深入,基因编辑技术的效率、准确性和安全性不断提升,动物精准基因编辑育种技术将逐渐向工程化和规模化方向发展,未来将在动物育种领域广泛应用,为动物新品种培育提供重要技术支撑。
基因编辑技术已成为动物育种的重要方法, 在动物育种领域得到十足的发展和应用。通过基因编辑技术精准创制了一批高产、优质、高繁殖和抗病育种新材料与新品系,如 MSTN 基因编辑的高产动物、抗繁殖与呼吸综合征的基因编辑猪和抗结核病奶牛等。家畜肉产量和品质、绒毛产量和品质等生产性状的改良对畜牧业高效可持续发展至关重要,通过基因编辑技术对关键基因的精准修饰已成为家畜重要经济性状改良的有效工具。利用基因编辑技术突变 BMP15 和 GDF9 基因, 显著提高了牛羊繁殖效率,创制了高繁殖动物育种材料。通过精确修饰调控肌肉生产的关键基因MSTN,获得的基因编辑猪、肉牛和羊均表现出双肌表型,产肉率显著提升。通过基因编辑技术将外源的 Fat1 基因成功插入猪基因组中,显著增加了猪肉中 n-3 多不饱和脂肪酸的含量,提高了猪肉的风味。精确突变 FGF5 基因,提高了高山美利奴羊的羊毛品质。针对动物抗病性状,发掘了一批宿主的重要抗病基因,通过精准改造病原进入细胞的受体基因,或者将抗性基因精准插入动物基因组安全性位点,培育多种特定病原抗性的动物育种新材料。使用 ZFNs 产生了具有抵抗非洲猪瘟能力的猪。通过基因编辑技术获得了抗猪瘟病毒(CSFV) 猪。使用 CRISPR-Cas9 敲除 CD163 受体而产生抗 PRRSV 的猪。针对导致猪伪狂犬病、猪繁殖与呼吸综合症、断奶仔猪多系统衰竭综合征等疾病的病原体,利用了 CRISPR-Cas9 系统对宿主进行了多个关键基因的修饰,培育了相应的抗病猪种。通过精确整合 Ipr1 和 NRAMP1 基因, 创制了抗结核病奶牛。使用 CRISPR-Cas9 技术对鸡 B 亚型肿瘤病毒基因(TVB)进行编辑,建立了抗 B 亚型白血病的鸡细胞系。以上研究表明, 动物精准基因编辑育种技术在动物新品种培育领域具有重要应用价值。
基因编辑技术解决了动物育种周期长等问题, 加速育种进程,降低育种成本。基于 CRISPR-Cas9 等系统的精准基因编辑技术可以运用到多种生物体和细胞的基因编辑中,且靶向修饰更精准、作用时间更短、操作更简单,获得的修饰后基因还可以实现种系遗传,在动物生产领域表现出巨大的潜力和优势。然而,当前的基因编辑技术仍处于研究和应用的初级阶段,还需要解决其脱靶效率高、编辑效率待进一步提高等关键问题。在未来的发展中,在实践中不断完善与成熟的基因编辑技术必将成为畜牧研究与生产领域的有力辅助工具,最终推动畜牧业的高质量发展。
“动物精准基因编辑育种技术”相关核心专利主要产出国家、主要产出机构分别见表 2.2.1 和表2.2.2 。核心专利公开量最多的是中国,为 3 860 项, 占比为 84.91%;排名第二的是美国,有 187 项专利, 占比为 4.11%,排名第三的是韩国,有 168 项专利, 占比为 3.70%。我国专利被引数比例为 56.63%,远超第二名的美国,但篇均被引频次只有 1.51,远远落后于法国、加拿大和美国。主要产出国家间的合作方面,美国、英国、荷兰、加拿大、澳大利亚之间有合作关系,其他国家间没有合作(图 2.2.1)。核心专利产出最多的机构是中国农业科学院,共有 237 项,中国农业大学和西北农林科技大学分别排在第二、第三名。被引数比例排名前三的机构分别是中国农业大学(4.35%)、中国农业科学院(3.26%)和华南农业大学(2.16%)。平均被引数最高的机构是中国农业大学,达到 3.07 次。而各主要产出机构间没有合作。图 2.2.2 为“动物精准基因编辑育种技术”工程开发前沿的发展路线。
《表 2.2.1》
表 2.2.1“动物精准基因编辑育种技术”工程开发前沿中核心专利的主要产出国家
| 序号 | 国家 | 公开量 | 公开量比例 /% | 被引数 | 被引数比例 /% | 平均被引数 |
| 1 | 中国 | 3 860 | 84.91 | 5 830 | 56.63 | 1.51 |
| 2 | 美国 | 187 | 4.11 | 2 854 | 27.72 | 15.26 |
| 3 | 韩国 | 168 | 3.7 | 155 | 1.51 | 0.92 |
| 4 | 俄罗斯 | 88 | 1.94 | 55 | 0.53 | 0.62 |
| 5 | 日本 | 69 | 1.52 | 190 | 1.85 | 2.75 |
| 6 | 法国 | 20 | 0.44 | 664 | 6.45 | 33.2 |
| 7 | 英国 | 20 | 0.44 | 480 | 4.66 | 24 |
| 8 | 澳大利亚 | 18 | 0.4 | 98 | 0.95 | 5.44 |
| 9 | 加拿大 | 13 | 0.29 | 382 | 3.71 | 29.38 |
| 10 | 荷兰 | 12 | 0.26 | 52 | 0.51 | 4.33 |
《表 2.2.2》
表 2.2.2“动物精准基因编辑育种技术”工程开发前沿中核心专利的主要产出机构
| 序号 | 机构 | 公开量 | 公开量比例 /% | 被引数 | 被引数比例 /% | 平均被引数 |
| 1 | 中国农业科学院 | 237 | 5.21 | 336 | 3.26 | 1.42 |
| 2 | 中国农业大学 | 146 | 3.21 | 448 | 4.35 | 3.07 |
| 3 | 西北农林科技大学 | 124 | 2.73 | 197 | 1.91 | 1.59 |
| 4 | 华南农业大学 | 123 | 2.71 | 222 | 2.16 | 1.8 |
| 5 | 扬州大学 | 96 | 2.11 | 114 | 1.11 | 1.19 |
| 6 | 华中农业大学 | 51 | 1.12 | 98 | 0.95 | 1.92 |
| 7 | 山东农业大学 | 45 | 0.99 | 72 | 0.7 | 1.6 |
| 8 | 四川农业大学 | 43 | 0.95 | 94 | 0.91 | 2.19 |
| 9 | 贵州大学 | 36 | 0.79 | 37 | 0.36 | 1.03 |
| 10 | 南京农业大学 | 33 | 0.73 | 54 | 0.52 | 1.64 |
《图 2.2.1》
图 2.2.1 “动物精准基因编辑育种技术”工程开发前沿主要国家间的合作网络
《图 2.2.2》

图 2.2.2 “动物精准基因编辑育种技术”工程开发前沿的发展路线
2.2.2 园艺作物基因编辑技术应用
园艺作物基因编辑技术主要是通过敲除几个碱基对或一段 DNA 序列从而改变原有基因序列的技术。基因编辑技术自问世以来已经发展更新 3 代: ZFNs 编辑技术、TALENs 编辑技术和 CRISPR- Cas9 编辑技术。其中 CRISPR-Cas9 基因编辑系统由于编辑效率高、操作简单、成本低等优点,成为目前园艺作物基因编辑的主要技术。基因编辑技术由最先对目标基因的剪切断裂引入突变,到现在可以实现对基因单个碱基的精准替换,以及大片段的精确插入或者替换等操作,技术上已经实现了重大突破。近年来,在番茄、西瓜、黄瓜、大白菜、柑橘、苹果、猕猴桃、月季、中国莲等重要园艺作物基因组解析方面取得突破性进展,在重要园艺作物营养和风味品质、产品器官形成、植株生长发育等重要性状的基因挖掘、园艺作物与环境互作机制等方面取得了一些重要成果,基因编辑技术起到重要推动作用。随着基因编辑技术的效率、准确性和安全性不断提升,园艺作物基因编辑技术将逐渐向工程化和规模化方向发展,未来将在园艺作物生产领域广泛应用,为培育更加优良的园艺品种及精准调控产量与品质奠定了良好基础。
基因编辑技术已成为园艺作物重要基因功能研究重要方法,在园艺作物研究领域得到长足的发展和应用。CRISPR-Cas9 系统首次于 2014 年在番茄中应用,研究人员成功将 Argonaute 7 基因敲除后使番茄叶片产生 Wiry 表型。随着 CRISPR-Cas9 技术的发展,在越来越多的园艺作物(包括黄瓜、草莓、香蕉、葡萄、苹果、西瓜和猕猴桃)中得到应用,在研究植物生长发育、产品品质、生物与非生物胁迫响应以及作物驯化等方面的重要基因功能方面取得了重要突破。例如,利用 CRISPR-Cas9 技术敲除草莓生长素响应因子 8(ARF8),产生了幼苗生长更快的植株;诱导番茄 SlCLV3 启动子突变, 获得了具有较多心室和较大果实的番茄;对抗番茄黄叶卷曲病毒进行基因编辑,获得比野生型番茄更高效的病毒抗性,积累的病毒基因组 DNA 较少, 且这种免疫活性可以世代传递;基因编辑技术通过引入远缘模式植物的已知农艺性状来加速各种野生植物的驯化过程。另外,基因编辑的有效性和目的性、无外源 DNA 的再生遗传材料和遗传改良作物监管法规的开放正在促进园艺作物中的基因编辑的研究与应用。
园艺作物基因编辑技术不仅在科学研究中起到巨大的推动作用,在商业应用方面,2021 年,一种经过 CRISPR 技术改造的番茄富含在日本上市销售,主要是降低了 γ- 氨基丁酸降解酶活性,因此这种番茄所含的 γ- 氨基丁酸是普通番茄的 5 倍, 这对 CRISPR 技术用于园艺作物育种来说是一个非常重要的里程碑。然而,仍有大部分园艺作物尚未有建立起成功的CRISPR 基因编辑体系,转化率低, 或无法得到基因编辑植株。因此,突破不同园艺作物的CRISPR 基因编辑体系,确定合理的目的性状、阐释性状的功能基因、提高编辑效率和分离无性繁殖作物中基因编辑引入的外源 DNA 等仍然是面临的挑战。
“园艺作物基因编辑技术应用”相关核心专利的主要产出国家、主要产出机构分别见表 2.2.3 和表 2.2.4。在核心专利数量方面,总体偏少,核心专利公开最多的国家是中国,为 38 项,占比为
《表 2.2.3》
表 2.2.3 “园艺作物基因编辑技术应用”工程开发前沿中核心专利的主要产出国家
| 序号 | 国家 | 公开量 | 公开量比例 /% | 被引数 | 被引数比例 /% | 平均被引数 |
| 1 | 中国 | 38 | 76 | 117 | 96.69 | 3.08 |
| 2 | 韩国 | 4 | 8 | 0 | 0 | 0 |
| 3 | 日本 | 3 | 6 | 1 | 0.83 | 0.33 |
| 4 | 德国 | 1 | 2 | 2 | 1.65 | 2 |
| 5 | 美国 | 1 | 2 | 1 | 0.83 | 1 |
| 6 | 瑞士 | 1 | 2 | 0 | 0 | 0 |
| 7 | 荷兰 | 1 | 2 | 0 | 0 | 0 |
| 8 | 俄罗斯 | 1 | 2 | 0 | 0 | 0 |
《表 2.2.4》
表 2.2.4 “园艺作物基因编辑技术应用”工程开发前沿中核心专利的主要产出机构
| 序号 | 机构 | 公开量 | 公开量比例 /% | 被引数 | 被引数比例 /% | 平均被引数 |
| 1 | 江苏省农业科学院 | 6 | 12 | 36 | 29.75 | 6 |
| 2 | 北京市农林科学院 | 4 | 8 | 2 | 1.65 | 0.5 |
| 3 | 新疆农业科学院园艺作物研究所 | 3 | 6 | 17 | 14.05 | 5.67 |
| 4 | 浙江省农业科学院 | 2 | 4 | 23 | 19.01 | 11.5 |
| 5 | 西北农林科技大学 | 2 | 4 | 6 | 4.96 | 3 |
| 6 | 浙江大学 | 2 | 4 | 2 | 1.65 | 1 |
| 7 | 中国农业科学院 | 2 | 4 | 1 | 0.83 | 0.5 |
| 8 | 南京农业大学 | 2 | 4 | 0 | 0 | 0 |
| 9 | 中国科学院上海生命科学研究院 | 1 | 2 | 14 | 11.57 | 14 |
| 10 | 华南农业大学 | 1 | 2 | 6 | 4.96 | 6 |
76.00%;排名第二的是韩国,有 4 项专利,占比为 8.00%;排名第三的是日本,有 3 项专利,占比为 6.00%。我国专利被引数为 117 次,比例为96.69%,远超第二名的韩国。各主要产出国家间没有合作。
核心专利产出最多的机构是江苏省农业科学院,共有 6 项,北京市农林科学院、新疆农业科学院园艺作物研究所分别排在第二、第三名。被引数比例排名前三的机构分别是江苏省农业科学院(29.75%)、浙江省农业科学院(19.01%)和新疆农业科学院园艺作物研究所(14.05%)。平均被引数最高的机构是中国科学院上海生命科学研究院,达 14 次。各主要机构间没有合作。
图 2.2.3 为“园艺作物基因编辑技术应用”工程开发前沿的发展路线。
2.2.3 林木全基因组选择育种
林木重要性状多为复杂的数量性状、遗传杂合性高,常规育种技术难以高效快速定向培育林木良种。分子标记辅助选择不能捕获微效位点、遗传作图分辨率低、数量性状遗传定位结果间无法互相验证,导致林木育种进展缓慢,人工林的生长和生产效率远不能满足经济和生态的需求。鉴于性状表型选择和分子标记辅助选择育种策略的不足, Meuwissen 等提出了基因组选择(GS),亦称为全基因组选择。GS 是一种全基因组范围的标记辅助选择方法,主要利用覆盖全基因组的高密度遗传标记信息,估计个体基因组范围的育种值 GEBV,并以 GEBV 为依据选择优良基因型。GS 是结合功能基因组研究成果和单核苷酸多态性(SNP)芯片技术建立的早期选择技术,基于林木种子或幼苗基因型,预测林木性状表现,准确鉴定并筛选带有优良基因的个体。为最终建立以 GS 为核心的林木智能设计育种体系,需要:研发GS 技术在林木亲本选配、杂交组合预测和优良子代早期选育的实现路径;精准挑选聚合多基因、表型优良的林木新品,科学控制育种过程;促进林木 GS 和基因编辑、QTL、全基因组关联分析(genome wide association study, GWAS)、表型组学多技术和学科的交叉融合,推动林木复杂性状关键育种基因的挖掘。
GS 的优势主要体现在如下几个方面。① 育种速度更快。相比表型选择,依据 GEBV 能进行早期个体选择,效率更高,能有效降低世代间隔,对育种周期长的林木育种最为关键。② 选择强度更大。实现高通量的基因分型比表型测定成本更低, 依据基因型能评估更多候选者,增大选育群体规模对子代群体庞大的植物育种尤为重要。③ 预测准确性高。基于分子标记的 GEBV 比单纯基于表型和谱系的 EBV 估算更准确,能极大地提高对微效位点的捕获功效,从而提高复杂数量性状预测的准
《图 2.2.3》
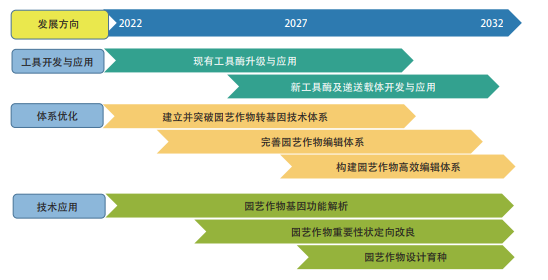
图 2.2.3 “园艺作物基因编辑技术应用”工程开发前沿的发展路线
确性和遗传标准差。④ 更有利于保持遗传多样性。通过有效整合不同双亲群体和育种项目的遗传材料,在大规模群体中开展遗传评估和选择,有利于保存丰富的遗传资源。⑤ 更节省时间和人力成本。能省去或减少子代测定工作,可直接对预选个体进行无性系化和子代测定试验。综上,GS 特别适合于育种周期较长、目标性状遗传复杂、表型测量难度大或成本高的林木树种,可显著提升目标性状的遗传改良精度,降低操作成本,是具有巨大潜力的林木育种策略。
GS 是目前动物和作物遗传育种的关键技术和研究热点,已在一些动植物的遗传改良工作中取得了重要进展,然而林木 GS 育种相对落后,尚处于初级阶段。自 2010 年以来,林木 GS 领域有近 80 项成果发表,多集中于桉树、云杉、火炬松、花旗松等用材树种,主要针对林木生长、材性和抗病虫等性状构建 GS 预测模型,并探究其影响因素,包括训练群体规模与组成、性状遗传率、标记数量、模型算法优化对比等。林木 GS 育种应用实例和跨越多世代的 GS 研究鲜有报道,林木 GS 育种滞后的原因包括:大多数树种遗传资源收集和种质评价工作不深入,缺乏用于快速建立大规模训练群体的已建成的试验林;遗传研究基础薄弱、高通量表型测定技术落后、标准化低成本基因型分型技术缺乏; 基因型 × 年龄 / 环境互作模型缺乏、针对林木特点分析工具缺少;树种参考基因组和分子标记质量低,会降低模型预测精度。
为克服困难促进林木 GS 育种发展,应在以下几个方向开展研究。① 建立统一基因型和表型获取平台:利用高密度 SNP 芯片等技术高通量、低成本、标准化采集基因型数据,通过高通量表型测定平台非破坏性、大尺度获取多维表型组数据。② 优化和开发林木 GS 模型算法:根据林木特点, 结合多组学、多环境和多年份数据,利用机器学习与深度学习,开发基因型× 环境/ 年龄互作模型, 实现 GS 算法的优化和提升,不断扩充和更新训练群体优化预测模型,提高未知气候环境中林木 GS 模型的普适性和长期功效性。③ 整合分析跨越多群体的上千株样本数据,综合 GS 和 meta-QTL 获得稳定可靠分子标记,开发特定性状选育 SNP 芯片产品。④ 利用 C++、Julia 和 R 等编程语言开发针对林木特点(多年生、异交和样本量庞大)的分析工具,最常用的R 语言分析速度慢,需开发能快速处理海量分子标记数据(>100 k)的分析软件。⑤ 对于难以获得高质量基因组的针叶树和多倍体物种,开展林木泛基因组研究,建立高质量参考基因组,以提高 GS 模型预测准确性。另外,林木GS 样本需求量大、成本高,应促进数据收集和共享, 注重研究机构和育种公司合作,保证长期稳定投入和研究。
“林木全基因组选择育种”工程开发前沿包含 2 项核心专利(表 2.2.4、表 2.2.5),共被引 7 次,均为中国专利,均来自华南农业大学,发明人是以林元震为核心的林木遗传育种研究团队。其中,专利“基于多性状的基因组选择进行林木多性状聚合育种”方法联合基因组选择技术与多
《表 2.2.4》
表 2.2.4“林木全基因组选择育种”工程开发前沿中核心专利的主要产出国家
| 序号 | 国家 公开量 公开量比例 /% 被引频次 | 被引频次比例 /% | 平均被引频次 |
| 1 | 中国 2 100.00 7 | 100 | 3.5 |
《表 2.2.5》
表 2.2.5“林木全基因组选择育种”工程开发前沿中核心专利的主要产出机构
| 序号 | 机构 公开量 公开量比例 /% 被引频次 | 被引频次比例 /% | 平均被引频次 |
| 1 | 华南农业大学 2 100.00 7 | 100 | 3.5 |
性状聚合技术,建立林木多性状聚合育种新方法体系,实现多性状定向、精确育种,显著缩短林木育种周期;专利“基于个体遗传竞争与环境空间分析的林木基因组选择”方法充分考虑遗传材料和种植环境的背景差异,采用 SNP 标记构建基因组关系矩阵,US 结构拟合个体遗传竞争效应, AR1 结构拟合环境空间效应,进行基因组选择分析,显著提高林木基因组选择的准确性。图 2.2.4 为“林木全基因组选择育种”工程开发前沿的发展路线。
《图 2.2.4》
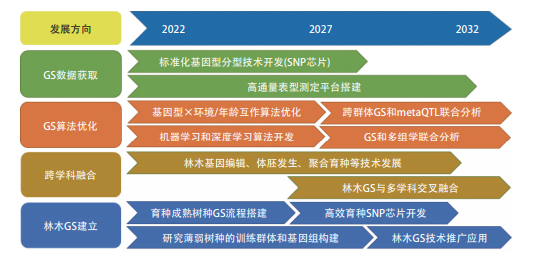
图 2.2.4 “林木全基因组选择育种”工程开发前沿的发展路线
领域课题组成员
课题组组长:
张福锁
专家组:
曹光乔 陈源泉 戴景瑞 韩丹丹 韩建永
韩 军 郝智慧 康绍忠 李德发 李道亮
李 虎 刘少军 刘平黄 李天来 刘晓娜
罗锡文 董朝斌 蒲 娟 齐明芳 申建波
沈建忠 王桂荣 王红亮 王军军 魏海燕
吴孔明 吴普特 武振龙 张福锁 张洪程
张守攻 张小兰 张 涌 赵春江 臧 英
周 磊 周 毅 朱齐超 朱旺升 朱作峰
课题组:
初晓一 郜向荣 李红军 李云舟 刘德俊
刘 军 师丽娟 孙会军 汤陈宸 王桂荣
姚银坤 张晋宁 赵 杰 周丽英
执笔组:
董朝斌 高 辉 韩丹丹 韩建永 胡 炼
金诚谦 李少锋 李 思 刘 军 刘晓娜
罗锡文 钱永强 钱震杰 权富生 孙康泰
孙世坤 田 静 王军辉 王军军 武振龙
邢志鹏 杨 青 赵春江 张 涌 张苗苗
周焕斌 周 毅 朱齐超











 京公网安备 11010502051620号
京公网安备 11010502051620号




