

《1. 从除虫菊到拟除虫菊酯——绿色化学的开端》
1. 从除虫菊到拟除虫菊酯——绿色化学的开端
《1.1. 化学过程》
1.1. 化学过程
除虫菊是从菊科植物灰菊头中提取出的6种天然亲脂性除虫菊酯的混合物,可以说是最成功、最有效的天然植物源性杀虫剂。除虫菊的杀虫特性自中世纪以来就已为人所知,并被广泛用于保护人们免受跳蚤、虱子和蜱虫的侵害,这些寄生虫传播了各种人类疾病,已造成数百万人死亡。然而,由于天然的除虫菊酯在空气、光线、水和土壤中会被迅速分解为无杀虫活性的产物,因此其在很大程度上局限于室内或个人使用。在20世纪 60~70年代,人们采取了一种系统的方法来改变天然除虫菊酯的光不稳定结构,同时保留了其快速且有效的杀虫作用和对哺乳动物低毒性等特性。最重要的是用卤素(如溴、氯和氟)取代天然酯酸基化的乙烯基部分的甲基,以及用光稳定性醇(如3-苯氧苄醇)取代天然醇部分。这些研究促使20多种合成除虫菊酯(拟除虫菊酯)被合成和商业化,并被广泛而大量地用于昆虫控制的各个方面,包括农业、病媒控制、家庭和宠物杀虫剂配制等,几乎人人都有可能接触到[1]。最成功的两种拟除虫菊酯是I型拟除虫菊酯,即氯菊酯(3-苯氧苄醇的亚甲基碳上没有α-氰基)和II型拟除虫菊酯,即溴氰菊酯(有α-氰基)。
《1.2. 作用位置和模式》
1.2. 作用位置和模式
天然除虫菊酯、合成拟除虫菊酯和 p,p ′ -二氯二苯三氯乙烷(DDT)在神经组织中有一个共同的主要作用靶点:电压门控钠离子通道(VSSC)。以上这些杀虫剂都会导致VSSC失活速度减慢和失活时间延长,最终导致钠离子内流增加和膜去极化。对于I型拟除虫菊酯,如氯菊酯,去极化导致后电位去极化,最终导致神经细胞重复放电和中毒性震颤综合征(T综合征)。在II型拟除虫菊酯(如溴氰菊酯)的情况下,失活被大大延长,导致大量膜去极化,并造成伴有中毒性唾液分泌综合征(CS综合征)的舞蹈手足徐动症。
《1.3. 《食品质量保护法》和美国国家环境保护局 / 风险杯分析》
1.3. 《食品质量保护法》和美国国家环境保护局 / 风险杯分析
1996年,美国国会通过了《食品质量保护法》(FQPA),并指示美国国家环境保护局(USEPA)从总暴露量和累积风险方面重新评估农药的允许量。如 FQPA所述,应增加与具有共同作用机制的杀虫剂相关的风险,如果总风险超过设定的量(风险杯),此类农药将被禁止使用。由于拟除虫菊酯在VSSC中具有共同的作用机制,并被广泛和集中地使用,因此USEPA开始了一项数据审查。2010年,USEPA通知注册人,在与人类健康风险评估关联更密切的低剂量条件下,测试未能解决青少年与成人对拟除虫菊酯敏感性可能存在年龄相关差异的问题。作为回应,拟除虫菊酯人类风险评估促进委员会(CAPHRA)于2011年成立,其代表拟除虫菊酯和除虫菊酯杀虫剂登记人的利益,并解决以下问题:儿童(也就是刚学会吃饭的幼儿)是否比成人对拟除虫菊酯的神经毒性更敏感?
为了解决这一问题,CAPHRA开发了一个研究项目,系统地检查与拟除虫菊酯神经毒性相关的毒物动力学(即吸收、分配、新陈代谢、分泌)和毒效动力学(受体-配体相互作用建立效力和效能)的各个方面,并区分新生大鼠所描述的敏感性是否与婴儿和儿童有关。2011年,CAPHRA接受了神经膜小组的提议,该小组是由马萨诸塞大学阿姆赫斯特分校、萨乌瑞吉纳大学和精密生物测定公司的研究人员组成的一个联合会,旨在对拟除虫菊酯的潜在年龄相关的毒效动力学差异进行体内/体外评估。以下是通过改进基于注射大鼠脑神经膜的非洲爪蟾卵母细胞和高通量电生理技术的微移植试验来解决这一问题的创新体外方法的发展综述。
《2. 需要解决的问题和合理的解决方案》
2. 需要解决的问题和合理的解决方案
《2.1. 问题——没有所谓的 VSSC》
2.1. 问题——没有所谓的 VSSC
为了研究拟除虫菊酯等神经毒性杀虫剂对离子通道的影响,人们采用了多种电生理学方法,包括体外细胞记录、全细胞膜片钳以及通过向非洲爪蟾卵母细胞中注射互补RNA(cRNA)来实现克隆通道的异源表达[2]。电生理学允许快速收集与离子通道门控过程速度相匹配的数据,是直接测定神经毒物对通道开启和关闭过程动力学所起作用的首选方法。然而,很多最常用于检测天然通道活性的制剂是非神经性[3]和(或)非哺乳动物性的[4,5]。此外,电压敏感离子通道,包括VSSC,是由多种亚基型经过选择性剪接、RNA编辑及包括糖基化、甲基化和磷酸化在内的翻译后修饰等步骤,在哺乳动物体内组装而成的。这种组装过程的分歧在于,并没有所谓的VSSC,正相反,存在着大量不同的通道,具有独特的动力学和药理学特性。因此,人们通常并不知道组织中表达的通道亚型的实际组成,也不知道它们的表达如何随时间或位置而变化。因此,要想逐个地测定拟除虫菊酯等神经毒物对个体VSSC的影响,即便有可能成功,也是一项困难的任务。由于许多异源表达的通道没有经过基因组、转录组和翻译后修饰,其使用也会导致其他问题。
《2.2. 一种解决方案——将神经膜片段微移植到非洲爪蟾卵母细胞中》
2.2. 一种解决方案——将神经膜片段微移植到非洲爪蟾卵母细胞中
另一种方法是将完整生化系统的优点与电生理学的灵敏度和快速获取数据的特点相结合,使上述方案的许多问题最小化。现有文献[6–13]构建了一种方法,将神经膜片段(即神经膜)注射到非洲爪蟾的卵母细胞中。这种使用许多不同生物的神经片段的方法确实有效,而且能直接测量流经完整离子通道的离子流,这些离子通道在标准的二电极电压钳(TEVC)电生理学方法下,通过适当的亚基构造和翻译后修饰,嵌入其内源性脂质中。然而,该技术的主要问题之一是卵母细胞与神经膜片段的结合具有变异性[9,14]。
《3. 我们的方法——将神经膜注入非洲爪蟾卵母细胞》
3. 我们的方法——将神经膜注入非洲爪蟾卵母细胞
《3.1. 方法的优点与缺点》
3.1. 方法的优点与缺点
微移植技术是将含有天然脂质和蛋白质的大鼠脑神经膜片段植入非洲爪蟾卵母细胞的质膜中,使其发挥作用。本方法的优点包括:①表达的蛋白质以内源性状态存在,并被其天然脂质所包围,与大鼠脑内的状态相似;②有许多神经靶点包括电压敏感离子通道,可供研究;③注射后1~2 h可快速研究离子电流;④仅注射几纳克的组织就足以测量离子电流;⑤新鲜和冷冻的脑组织皆可使用;⑥该程序可以研究不同物种、年龄和大脑区域之间的差异;⑦可通过电子方式或药理学的方法分离特定的离子电流;⑧使用标准的电生理学方法可快速采集数据,并且适用于高通量系统(如Robocyte2®)[15]。这种方法的缺点有:①在许多组织和物种中存在非特异性蛋白;②由于多个靶蛋白的表达而产生复杂的离子电流;③神经片段结合的水平和方向可变[9]。
《3.2. 将神经膜注入卵母细胞的初试验》
3.2. 将神经膜注入卵母细胞的初试验
借助自动免疫印迹和TEVC电生理学技术,我们开始将幼年与成年大鼠脑组织片段微移植入非洲爪蟾光滑的卵母细胞中,来探究其年龄相关的表达模式、功能性差异以及对I型拟除虫菊酯(氯菊酯)和II型拟除虫菊酯(溴氰菊酯)的敏感性等方面的差异[16–18]。
经过这些初步研究,我们证实:成年大鼠脑神经膜微移植入卵母细胞的质膜,在天然态下这些细胞包含了在大鼠脑内发现的全部9种VSSC亚型,在膜去极化时诱发一种复杂的外向离子电流,其中一部分对河豚毒素(TTX,一种特殊的VSSC阻滞剂)很敏感,当缓冲液中的钠离子被非渗透的胆碱离子取代时,该离子流也随之消失[16]。此外,研究发现,去极化时所见的外向电流主要是由钙激活氯电流引起的,该电流可通过使用尼氟灭酸(NFA)进行实质性阻断,显示出TTX敏感的内向电流(图1)。
《图1》
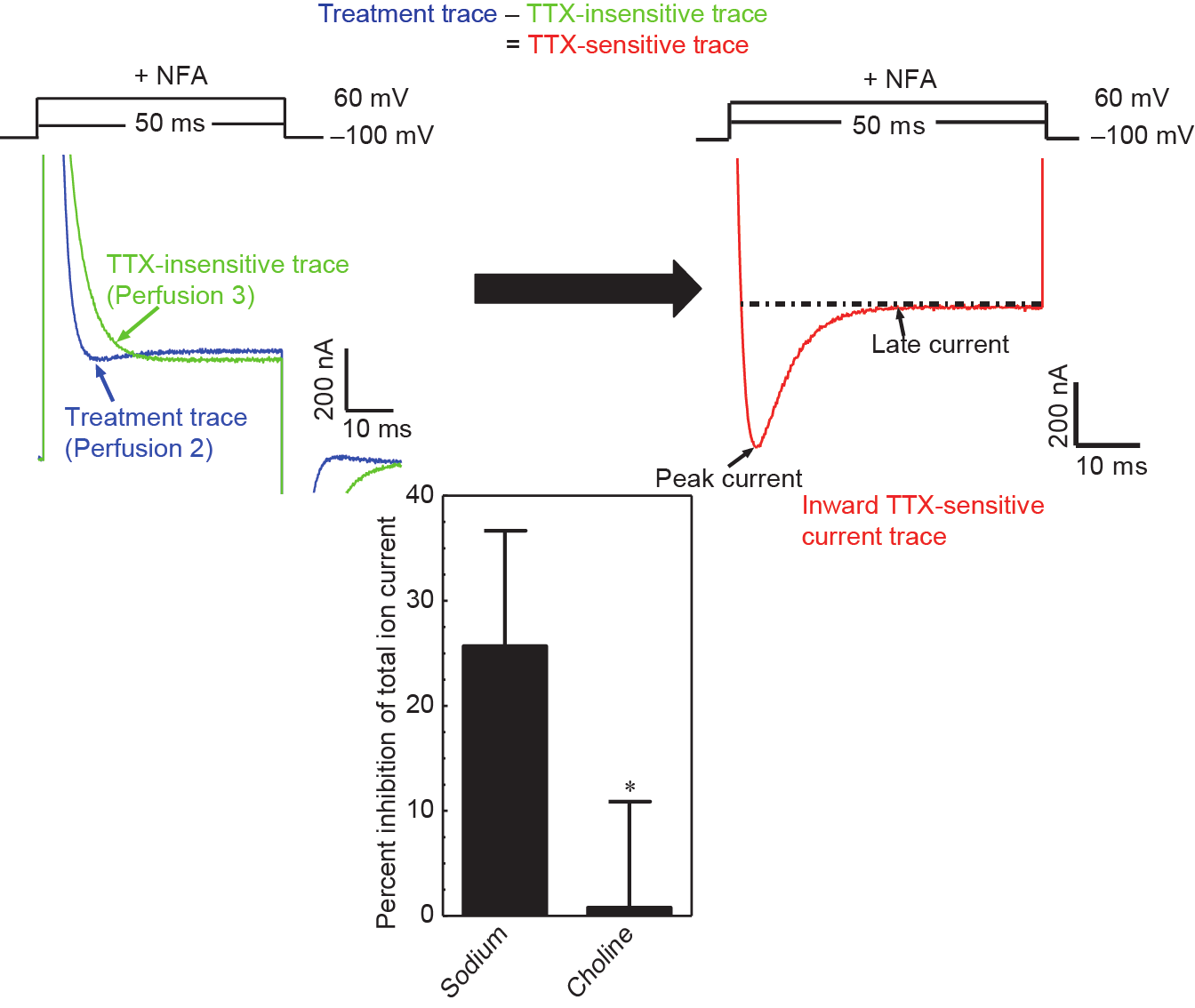
图1. 在NFA存在时,被微移植入非洲爪蟾卵母细胞的PND90大鼠脑神经膜组织中,由杀虫剂诱导出TTX敏感内向电流。TTX敏感内向电流(右端)是由处理轨迹和TTX不敏感轨迹(左端)之间的差异产生的。
这种TTX敏感内向电流在DDT [一种神经毒性杀虫剂,也作用于VSSC;但非神经毒性代谢物二氯二苯二氯乙烯(DDE,一种非神经毒性代谢物)除外]、氯菊酯和溴氰菊酯的协同作用下以浓度依赖性的方式增长,用脉冲去极化后曲线下面积(AUC)来测量[17,18]。尽管TTX敏感的AUC值同使用Hill方程生成的与拟除虫菊酯相关的S形浓度-响应曲线相当吻合,但由标准误差条判断的变异很大,降低了该方法在监管问题上的有效性(图2、图3)。
《图2》
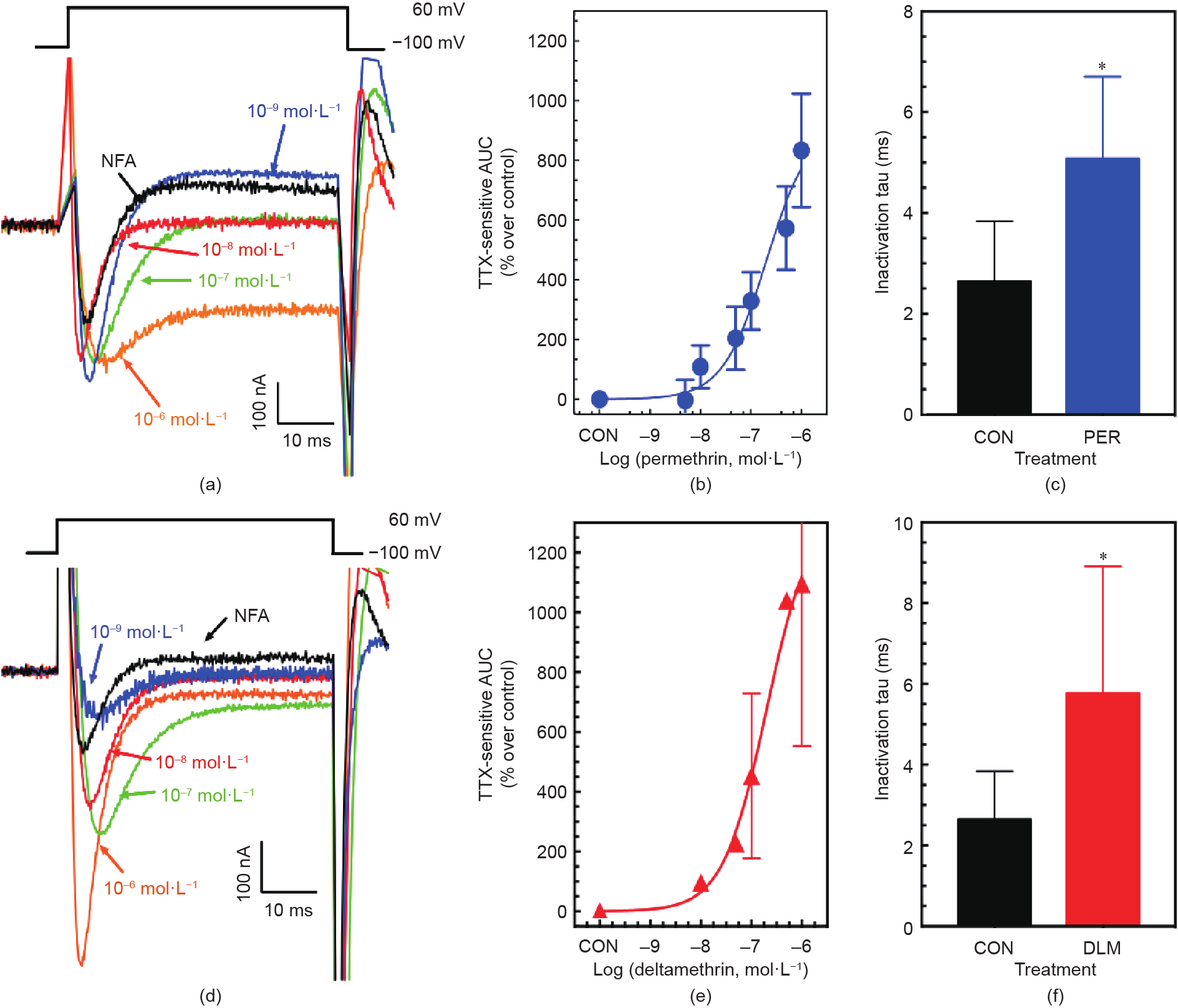
图2. 存在NFA(一种由Ca2+激活的氯离子通道阻滞剂)时,增加拟除虫菊酯浓度对PND90大鼠脑神经膜微移植到非洲爪蟾光滑的卵母细胞引起的去极化、TTX敏感内向电流的影响。(a)电生理的TTX敏感电流显示了氯菊酯浓度增加的影响;(b)浓度依赖响应曲线(CDRC)显示了氯氰菊酯对AUC电流轨迹的影响,其表现为超出对照的百分比;(c)10–6 mol·L–1的氯菊酯对失活τ值的影响数值(τ0.5,使50%的去极化VSSC失活所用的时间)的影响;(d)说明溴氰菊酯浓度增加影响的电生理TTX敏感电流轨迹;(e)CDRC显示了溴氰菊酯对AUC值的影响,AUC值表示为超出控制的百分比;(f)10–6 mol·L–1 的溴氰菊酯对失活 τ 值(τ0.5)的影响。TTX敏感内向电流是通过从总电流中减去TTX存在时的实验轨迹来确定的。超出控制的百分比值=[(处理AUC–NFA控制AUC)/ NFA控制AUC]×100。失活τ值是通过使用Origin(版本8.6,Origin实验室,美国马萨诸塞州北安普顿)将失活期间的后期电流轨迹拟合成指数衰减方程得到的。*表明,样本均值显著不同于对照组(NFA)卵母细胞(单样本的学生t检验, 10–6 mol·L–1: P< 0.05)。CON:对照。
《图3》
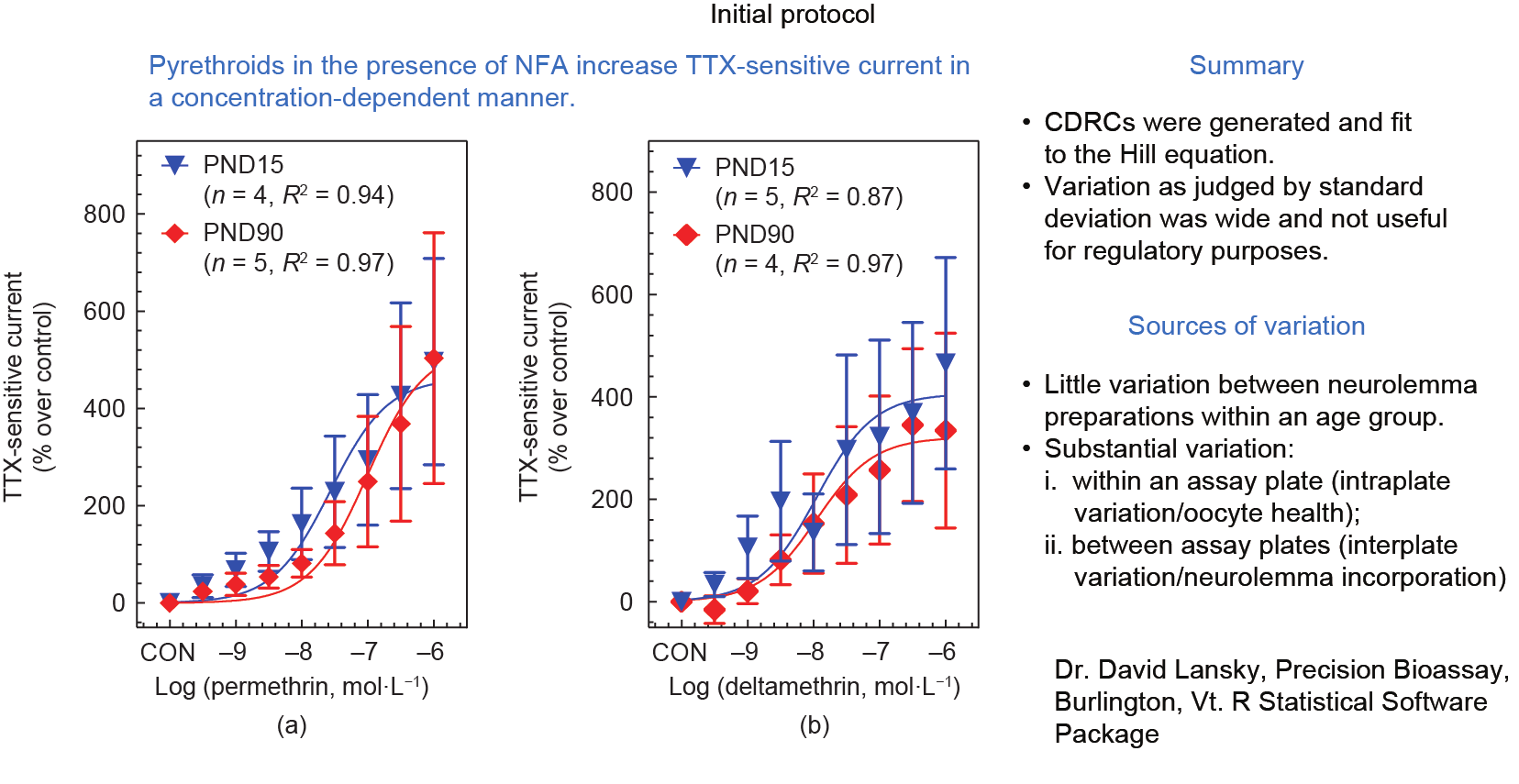
图3. 存在NFA时,增加氯菊酯(a)和溴甲烷(b)浓度对微移植到非洲爪蟾卵母细胞中的幼年PND15或成年PND90大鼠脑神经膜去极化诱发的 TTX敏感电流的影响。
《4. 提升精密度与分析性能》
4. 提升精密度与分析性能
通过建立这一程序作为毒理学相关的体外试验,对试验方案、数据采集标准和数据分析方法的创新变化进行收集,大大提高了精度,从而提高了分析性能[18]。在本文的其余部分中,我们系统地研究了这些改进并概述出一种可用于使用其他靶点和其他动物/组织的通用方法,以提高类似试验的性能。
《4.1. 对提高卵母细胞整体健康的检测水平的改变》
4.1. 对提高卵母细胞整体健康的检测水平的改变
为了改进神经膜微移植试验,通过将氯菊酯反应与线性混合模型拟合确定变异源,随机效应与神经膜制备、卵母细胞制备和单个卵母细胞有关。这些拟合的结果包括与每个来源(神经膜制剂、卵母细胞制剂和卵母细胞)相关的估计方差分量。结果表明,与神经膜制剂相关的方差分量比卵母细胞制剂和单个卵母细胞的方差分量小得多[18]。利用这些信息,我们进行了一个系统的评估:①板内单个蛙卵母细胞活力的变化(板内变化);②调整/最小化不同蛙卵母细胞组间质膜神经膜结合的变异性的程序(种间变异)。
根据原始方法使用的分析设置方案,很显然,卵母细胞的整体健康水平和活力随着完成完全浓度依赖性反应曲线(CDRC)的时间的推移而下降,导致只有 30%~50%的卵母细胞提供有用的数据(如TTX敏感的内向电流)。为了尽量减少卵母细胞健康状况的下降,对方案做了以下几点改变:手术切除卵母细胞后,只对 V期和VI期的健康卵母细胞进行去核和选择注射;在全分析微孔板中,96个卵母细胞在交错的时间点(每6 h)被注射0.5 mg·mL–1的神经膜,以确保更一致的结合时间;只在电生理试验的进行时间内记录放置在检测板上的缓冲液中的卵母细胞;以及将记录卵母细胞的时间间隔从15 min缩短到11 min。此外,在不同的微板之间交替使用拟除虫菊酯处理一排卵母细胞,奇数值浓度应用于第3~5行,偶数浓度应用于第6~8行。在下一次微板测定时将这一顺序颠倒,以此类推,从而使任何潜在的板块效应最小化。
《4.2. 数据采集标准的变化》
4.2. 数据采集标准的变化
建立了一个四步灌注的方法,对每个卵母细胞进行四次单独的电生理扫描,以在数据分析之前选择注射神经膜的卵母细胞,这些卵母细胞都具有TTX敏感的内向电流(TTX清洗方案)。先用1×ND96培养基和10-4 mol·L–1 NFA灌注卵母细胞2 min,然后进行电生理记录(扫描1)。然后用含有10-5 mol·L–1 TTX 的1×ND96培养基和10-4 mol·L–1 NFA灌注卵母细胞 2 min,并进行第二次记录(扫描2)。用1×ND96培养基和10-4 mol·L–1 NFA的灌注冲洗TTX 1 min。为了检验拟除虫菊酯处理的效果,将氯菊酯或溴氰菊酯[5 × 10-9 ~1 × 10-6 mol·L–1,置于1.0%二甲基亚砜(DMSO)中]和10-4 mol·L–1 NFA在1×ND96培养基中灌注5 min,并记录另一次电生理测量结果(扫描3)。最后,每个卵母细胞在含有10-4 mol·L–1 NFA和10-5 mol·L–1 TTX的 1×ND96培养基中再灌注2 min并记录(扫描4)。每一次灌注均通过单独的管道输送,使用隔离的自动阀门,以0.5 mL·min–1的速率进行。在四次灌注后,记录电流轨迹(扫描1~4),并进行漏减方案。漏减产生了反映灌注处理效果的单一电流轨迹。如前所述[16,19],该轨迹是从–100 mV保持电位到+60 mV持续50 ms的阶跃(脉冲)去极化的结果。如前所述[20],通过从总电流中减去TTX不敏感电流来确定TTX敏感的内向电流。为了确定在NFA存在下是否发现了“内向”TTX敏感电流,从扫描1中减去扫描2。选择产生TTX敏感内向电流在–100~4000 nA之间的卵母细胞(由AUC测量)用于数据分析。
这些板内方案的改变将可用于平板数据分析的卵母细胞的比例从30%~50%提高到80%以上,这反映了神经膜注入卵母细胞的整体健康状况的显著改善。虽然使用改善卵母细胞健康的卵母细胞选择标准和方案能够在分析板基础上产生较少的变异性,但所获得的数据仍然不够一致,不足以进行监管评估。
《4.3. 数据分析的变化减少了分析的可变异性》
4.3. 数据分析的变化减少了分析的可变异性
为减少在种间基础上的可变性,使用方程(1)将 AUC值转换为对照响应值的比例。

式中,VTR mean AUC是用70 µmol·L–1藜芦定处理的每个卵母细胞检测板的AUC平均值(VTR,一种持久的VSSC 激活剂,与拟除虫菊酯不同的位点结合并作为阳性对照); Control mean AUC是每个没有拟除虫菊酯或VTR处理的卵母细胞检测板获得的AUC的平均值(阴性对照);treatment mean AUC是在拟除虫菊酯处理的情况下,从扫描4中减去扫描3得到的AUC。
通过该过程,特异性反应范围由未经拟除虫菊酯或VTR处理的卵母细胞(阴性对照)和经70 µmol·L–1 VTR处理的卵母细胞(阳性对照,无拟除虫菊酯)的 AUC的平均值限定。在没有拟除虫菊酯处理的情况下,控制响应的比例预计为0。随着拟除虫菊酯浓度的增加,控制响应的比例随着拟除虫菊酯处理的卵母细胞AUC 的增加而增加,接近最大值1.0。当数据从0到1有界时,单个分析数据直接拟合到logit-log(广义)线性回归。
《5. 结论》
5. 结论
初步尝试将小鼠脑神经膜片段微移植到非洲爪蟾卵母细胞中,研究拟除虫菊酯杀虫剂对天然VSSC的影响,结果在NFA存在的情况下,检测到去极化依赖的、TTX 敏感的内向电流随着拟除虫菊酯浓度的增加而增加。这些创新的发现支持了这样的观点,即这项技术是一种和毒理学相关的有用的体外方法,用于研究神经毒素和毒物在其原状态下对其靶点的影响。尽管CDRC与生成它们的Hill方程完美地吻合,但由标准误差确定的变化很大,因而在监管情况下降低了它们的有用性。通过迭代方法,我们系统地检查了检测方案的变化,以提高卵母细胞的整体健康水平,并改进数据采集和数据分析,如表1所示。
《表1》
表1 减少神经膜注射卵母细胞试验的变异性的步骤,以获得随着拟除虫菊酯浓度增加而产生的更一致的CDRC

根据CDRC的标准误差和改进的总体R2 值(图4)来判断,这些修改的方法可使CDRC的变异性大大降低,并显著提高了分析精度,从而提高分析性能。利用这种改进的方法,我们目前正在评估拟除虫菊酯对幼年大鼠和成年大鼠脑神经膜的天然VSSC的毒性动力学的潜在年龄相关差异。
《图4》
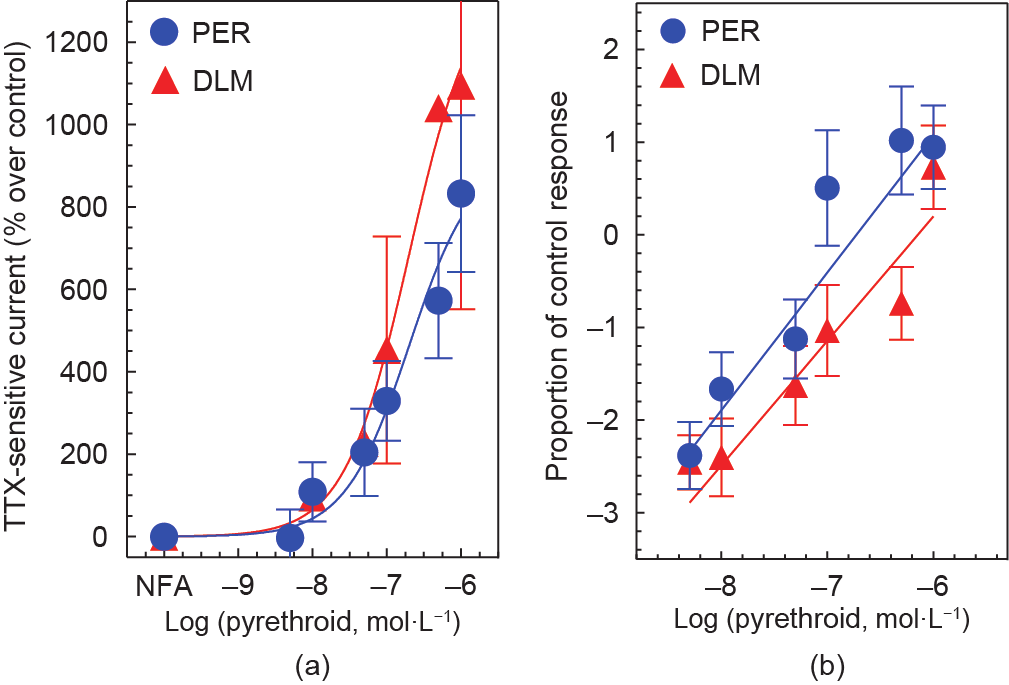
图4. NFA存在下,成年PND90大鼠脑神经膜微移植到非洲爪蟾卵母细胞中的拟除虫菊酯CDRC混合模型回归分析。(a)氯菊酯和溴氰菊酯的未调整数据集显示为超过对照反应的百分比;(b)使用分析改进获得的标准化数据集,以减少氯菊酯和溴氰菊酯的变异性,以对照反应所占比例的logit-log表示。
《6. 未来发展的前景》
6. 未来发展的前景
非洲爪蟾卵母细胞质膜片段微移植是一种体外方法,已被用于使用标准的电生理方法来研究天然态下的各种通道和受体[6–14]。令人遗憾的是,神经膜结合的变异限制了它在定性分析方面的应用。通过使用特定的阻断剂(如TTX和NFA),我们能够分离出一种去极化依赖的、TTX敏感的内向钠离子电流,这种电流是以浓度依赖的方式随着拟除虫菊酯和滴滴涕浓度的增加而增加,通过延长通道门控动力学的失活过程,模拟它们对卵母细胞中异源表达的单个VSSC的作用,从而验证了基于神经膜的方法。此外,将VTR(VSSC的一种特异性激活剂)加入阳性对照中,可以使不同批次卵母细胞之间的神经膜结合规范化,减少变异性,并促使该分析在定量方面有用。以这种方式,上述创新变化的集合概述了一种通用方法,可用于使用其他靶点和其他动物/ 组织提高类似试验的性能。重要的是,对于各种电压和配体门控的通道和受体,选择性激动剂和拮抗剂的可用性丰富且不断增加,这些通道和受体可如上所述用于提高分析性能。此外,已经证实该方法可以利用从尸体组织样本中获得的人脑碎片发挥作用[11];这允许直接比较人体靶位点上与非人类动物模型靶点上的毒物,从而验证了我们当前的动物模型的有用性。虽然目前这只是个推测,但有许多非神经组织(脂肪、肝脏、肌肉等)具有与神经组织相同的靶点[如VSSC、烟碱型乙酰胆碱受体、 -氨基丁酸(GABA)-氯离子通道、甘氨酸-氯离子通道等],它们都可以用上述分析方法进行研究。特别是,常见的环境污染物如多氟烷基物质(PFAS)和农药对这类非靶组织的作用尚未得到详细研究,尽管它们在代谢综合征中的作用(包括肥胖和2型糖尿病发病率的增加)已被广泛认可。
-氨基丁酸(GABA)-氯离子通道、甘氨酸-氯离子通道等],它们都可以用上述分析方法进行研究。特别是,常见的环境污染物如多氟烷基物质(PFAS)和农药对这类非靶组织的作用尚未得到详细研究,尽管它们在代谢综合征中的作用(包括肥胖和2型糖尿病发病率的增加)已被广泛认可。
《Acknowledgements》
Acknowledgements
This work was supported by the Council of the Advancement of Pyrethroid Human Risk Assessment (CAPHRA) (#S17110000000004).
《Compliance with ethics guidelines》
Compliance with ethics guidelines
The sponsor had no role in the study design, in the collection, analysis or interpretation of the data; in writing the article; or in the decision to submit the article for publication.
John Clark and Steve Symington declare that they have no conflict of interest or financial conflicts to disclose.











 京公网安备 11010502051620号
京公网安备 11010502051620号




