

《1、 引言》
1、 引言
随着材料科学的发展[1‒3],大量呈现多样类生命运动的人工驱动器被开发出来[4‒5],为仿生机器人带来了新的希望[6‒9]。鞭毛和纤毛是细胞的运动附属物,它们存在于诸多生物系统中,并实现着广泛的功能(如推进、进食和运输)[10‒12],这些引起了仿生学研究人员的极大关注。目前,已经揭示鞭毛和纤毛采用了截然不同的运动策略来支持微生物的生命活动[13]。通常,鞭毛通过手性波推动自身前进,表现为二维的平面波运动[14]和三维的螺旋波运动[15‒16];而纤毛通过二维或三维的不对称搏动来实现有效的运动[17‒18]。为了模仿这些运动,科学家们已经开发了许多不同的驱动机制来构造仿生驱动器,如磁驱动[19‒22]、电驱动[23‒26]、光驱动[27‒28]、超声驱动[29]和流体驱动等[30‒31]。基于这些仿生系统的研究已经惠及广泛的工程应用,包括但不限于货物操纵[32‒33]、液体操作[34]、低雷诺数环境推进[35]、微流控[36‒38]和医疗机器人[39‒40]等。
随着对低雷诺数流体动力学的了解,我们更深刻地理解了自然界中丰富的鞭毛/纤毛运动模式,并能通过适当的工程设计来复现它们,包括细长丝结构的平面摆动[41]、螺旋结构的旋转推进[42‒43],以及发状刺激响应驱动器的不对称搏动[26,28,44‒46]。然而,自然界中一个有趣的事实是,所有上述真核鞭毛/纤毛的运动(即鞭毛的平面/螺旋波运动和纤毛的二维/三维不对称搏动)都是从相同的9 + 2生物结构演化而来的[47‒50]。但由于缺乏统一的研究视角和对复杂生物内部结构与驱动机制的深刻理解,设计一个单独的工程结构来实现所有这些运动模式仍然是一个巨大的挑战。迄今为止,我们还不能完全复制大自然的智慧,即从同一结构出发,来演化出丰富的生物运动形式。这使仿生学研究无法从迷人的生物内部结构中获得任何灵感,也使我们难以为人工鞭毛和纤毛建立统一的仿生设计原理。
这项工作中,我们研究了鞭毛和纤毛的内部结构与驱动机制,发现根据动力蛋白的激活状态,它们所在的微管可以简化分类为三个功能区域,并通过特定的驱动策略产生不同的外部宏观运动。受此启发,我们提出了一种新的设计理念,通过杆嵌入铸造工艺制造了一个三通道的一体式管状驱动器。正如微生物由相同的9 + 2结构进化出丰富的运动那样,这一管状驱动器能够通过对每个通道的驱动编程来实现各类二维或三维的鞭毛和纤毛运动。此外,我们扩展了更多不对称的纤毛搏动模式,并通过低雷诺数推进测试进一步验证了它们的有效性。结合理论分析和实验验证,这项研究为构建鞭毛和纤毛启发的人工系统提供了新的见解,并展示了一个有力的、专注于底层生物学机制的仿生学案例。
《2、 材料和方法》
2、 材料和方法
《2.1 鞭毛/纤毛内在驱动机制的统一物理模型》
2.1 鞭毛/纤毛内在驱动机制的统一物理模型
如图1(a)所示,鞭毛和纤毛具有相同的、由九个双联体微管(DMT)和一个中心对复合体(CPC)组成的9 + 2结构[51]。在连接蛋白的限制下,动力蛋白在两个双联体微管之间的滑动最终导致了双联体微管的弯曲,见图1(b)中的(i)。依靠这种机制,鞭毛/纤毛的每一小节都可以独立地运动,且当双联体微管从根部(细胞头部)到尾部依次弯曲时,鞭毛/纤毛实现了最终的外部运动。为了阐明它们的内在驱动机制并获得统一化的原理,我们取连续鞭毛/纤毛的一个无穷小元素(定义为δl)进行分析。如图1(b)中的(ii),以δl中的双联体微管8号和9号为例,弯曲可以看作是由沿着动力蛋白的力引起的运动。假定每个动力蛋白具有相同的微元力(
(1)
式中,
(2)
式中,ψ和χ分别为力矢量与x轴正方向和y轴正方向的夹角;
《图1》

图1 设计原理。(a)9 + 2结构的示意图;(b)(i) 9 + 2结构中双联体微管弯曲机制的示意图;(ii)根据动力模型,横截面视角的鞭毛/纤毛微元的合力(如
这个统一模型表明鞭毛/纤毛的截面运动起主导作用,它们沿细长鞭毛/纤毛身体的传递(有相位差)决定了外部游泳模式。基于这样的事实,我们用所提出的统一模型研究了各种鞭毛/纤毛运动的驱动机制,发现它们都可以看作是由三个功能区域采用不同的驱动策略产生的(详见附录A中的图S1至图S4和补充文本)。对于鞭毛的平面波运动,两个双联体微管组(双联体微管2~4号,双联体微管7~9号)上的动力蛋白通过交替地激活和抑制实现弯曲运动,其水平运动分量受o-SUB5-6结构的限制[47]。因此如图1(c)所示,我们考虑将九根双联体微管划分为两个驱动区域(区域1和2)和一个限制区域(区域3)。两个驱动区的交替激活实现了鞭毛的摆动,在区域3的限制下,保证了其在准平面内运动。九根双联体微管上动力蛋白的顺序激活实现了鞭毛的螺旋波运动[48]。为简化这一过程,三个连续激活的双联体微管被视为一个驱动区域,那么就可看作三个独立驱动区域的顺序激活导致了最终的螺旋运动[图1(d)]。在二维纤毛搏动中,动力冲程和恢复冲程(纤毛搏动中一个完整运动周期的两个过程)分别由两个交替激活的驱动组(双联体微管1~4号和双联体微管6~8号)主导[49]。这里将9 + 2结构分为三个驱动区域,具体来说,有更多双联体微管的驱动组被视为包含两个同步激活/抑制的驱动区域,而另一个驱动组(有较少的双联体微管)则是第三个驱动区域[图1(e)]。至于三维纤毛搏动,两个双联体微管组(双联体微管2~4号,双联体微管6~8号)交替激活,它们分别支配动力冲程和恢复冲程[50],其他双联体微管上的动力蛋白未被激活。因此,我们考虑将九根双联体微管分成两个驱动区域和一个非活动区域[图1(f)]。上述分析表明,不同策略驱动下的三个功能区域可以产生丰富的鞭毛/纤毛运动形式[图1(g)]。
《2.2 三通道管状驱动器的制作与校准》
2.2 三通道管状驱动器的制作与校准
为展示生物学机制及所提出的仿生概念,我们制作了一个具有三个独立可寻址通道的一体式管状驱动器,以复现上述的鞭毛和纤毛运动。如图2(a)所示,软驱动器由硅橡胶(Ecoflex 00-30,美国Smooth-On公司)通过杆嵌入铸造工艺制成。在预处理过程中,将Ecoflex-A和Ecoflex-B按1∶1的比例混合,在真空条件下搅拌、脱气后得到均质的复合弹性体。然后,将混合液体注入嵌入了三个金属棒(直径约3 mm)的3D打印五件式模具中。将模具置于空气中12 h后,通过移除3D打印的外壳和金属棒,将固化的三通道软驱动器脱模。最后,驱动器的尖端(P点)用新的未固化的Ecoflex 00-30密封(厚度约2 mm),另一端的三个通道(I、II和III)分别连接到入气管,并用未固化的Ecoflex 00-30密封。制作的驱动器长65 mm,具有三个相同的通道(直径约3 mm),尺寸参数如图2(b)所示,具体数值参见附录A中的表S1。
《图2》

图2 一体式三通道管状驱动器的制作、驱动用实验装置和校准。(a)驱动器的制作说明:包括预处理、成型和脱模步骤;(b)三通道管状驱动器及其尺寸参数图示;(c)驱动器模拟生物运动的实验装置示意图;(d)~(f)驱动器弯曲运动校准(单通道充气、双通道充气和三通道充气)。
管状驱动器由气动泵系统驱动[详见图2(c)和附录A中的补充文本]。与9 + 2结构中的双联体微管一样,驱动器的三个通道相互平行连接(即为一体化结构)。当空气沿软通道注入时,会因压力失衡而产生横向弯曲,这与微管在动力蛋白运动和固定连接蛋白限制下产生的被动弯曲非常相似。这种与真实生物结构本质相似的仿生驱动器将用于呈现本文中不同鞭毛和纤毛的截面运动。
我们通过单通道、双通道和三通道充气来校准管状驱动器的弯曲性能。为了定量地描述弯曲性能,定义了变形长度(δ1、δ2和δL,其中,δ1、δ2是水平变形长度;δL是垂直变形长度)和偏转角(θ1和θ2)对三种类型做预测试,并得到这些参数在不同充气量(z)下的变化。如图2(d)~(f)所示,当充气体积从0到2.5 mL变化时,上述参数的变化范围如下:δ1: 0~43.3 mm, δ2: 0~34.0 mm, δL: 0~7.2 mm, θ1: 0~73o, θ2: 0~68o。预实验也表明了对于不同运动模式的验证和演示而言,相对运动误差可以忽略不计。值得注意的是,这里的误差主要来自两个方面:驱动器的制造误差和气动泵系统的机械误差。采用精细的制造工艺和高精度的系统,将有效提高运动精度。特别是当仿生系统用于微型操作设备(如微流体系统)时,精确运动控制和稳定性的要求尤为重要。除了气驱动之外,所提出的设计概念也可以通过其他驱动机制来实现。具体而言,合适的驱动器被期望是一体化的,且具有独立可控的内部响应组件,这些响应组件的协同可实现独特且多变的外部运动。例如,基于液晶弹性体(LCE)和液压驱动的人工纤毛驱动器就非常符合这些要求[27,45]。我们的设计原理和驱动策略有望应用于具有上述特征的仿生系统,并为其带来丰富的运动模式。关于一体式管状驱动器的其他细节可以在附录A中的图S5和补充文本中找到,包括驱动器的运动学分析、有限元法模拟(FEM)和轨迹跟踪方法。
《3、 结果和讨论》
3、 结果和讨论
《3.1 自然界中二维/三维鞭毛/纤毛运动的再现》
3.1 自然界中二维/三维鞭毛/纤毛运动的再现
三通道管状驱动器可以再现自然界中各类鞭毛和纤毛运动(包括鞭毛的平面/螺旋波运动和纤毛的二维/三维搏动)。就鞭毛的平面波运动而言,我们交替地对通道I和II进行充气/放气,充气体积范围从0到λ(峰值λ = 2.5 mL),同时保持通道III同步为通道I和II提供限制以纠正弯曲方向[图3(a)]。通过提供合适的充气比率(通道I/II∶通道III = 4∶3,详见图S6和附录A中的补充文本),得到了与鞭毛平面波运动中横截面视角运动一致的驱动器平面弯曲轨迹[图3(b)和附录A中的视频S1]。另一方面,对于鞭毛的螺旋波运动,我们依次交替地向三个独立的通道(I、II和III)充气,并保证相同的驱动相位差(2π/3)和峰值体积(λ),见图3(c)。这时驱动器呈现出近圆形的轨迹,很好地再现了鞭毛螺旋波运动的横截面视角运动[图3(d)、(e)和附录A中的视频S1]。这里我们注意到两类鞭毛的横截面视角运动都是可回溯的,因此它们不能独立提供有效的推进。然而,就像自然界中观察到的真实生物运动一样,这种截面运动可以沿着细长体传递,从而形成有效的二维或三维手性推进波(由多周期截面运动构成)。
《图3》

图3 通过一体式驱动器执行的鞭毛截面运动模拟。(a)平面波运动的示意图和充气驱动策略;(b)平面波的截面运动沿着驱动器形成了面内搏动;(c)螺旋波运动的示意图和充气驱动策略;(d)螺旋波的截面运动沿着驱动器在三维空间中形成近圆形路径;(e)不同时间下记录的螺旋波运动轨迹(含俯视图和正视图)。t:时间;T:一个完整运动周期的时间。
二维纤毛搏动的完整运动周期包括动力冲程和具有不同路径的恢复冲程(即路径不对称)[18]。按图4(a)中的驱动策略,我们首先向通道III中充入λ体积的气体使其膨胀弯曲,作为初始状态,见图4(b)。在动力冲程中,向通道I和II同步充入λ体积的气体(阶段1:0~T/4,其中,T是一个完整运动周期的时间),然后将通道III中的气体完全放出(阶段2:T/4~T/2),便完成了动力冲程。在恢复冲程中,将通道I和II中的气体排出(阶段1:T/2~3T/4),然后通道III重新充入λ体积的气体(阶段2:3T/4~T)以完成恢复冲程。如图4(c)所示的尖端轨迹和附录A中的视频S1,三通道管状驱动器很好地展示了具有路径不对称特征的二维纤毛搏动。在三维纤毛搏动中,围绕着轨迹的平均轴偏离表面法线,呈现方向不对称特征[17]。为了复现这种三维搏动,我们交替地对通道I和II进行充气/放气,同时保持通道III不充气[图4(d)]。如图4(e)所示,这种情况下的驱动器在俯视图中呈现出相对于底部中心B点的偏心轨迹[图4(f)和附录A中的视频S1],很好地再现了三维纤毛搏动的方向不对称性。与鞭毛的手性波不同,二维和三维纤毛搏动的截面运动是不可回溯的,这也与自然界纤毛中观察到的不对称运动特征一致。
《图4》

图4 通过一体式驱动器执行的二维和三维纤毛搏动。(a)二维纤毛搏动的示意图和充气驱动策略;(b)其截面运动沿驱动器形成具有路径不对称的平面内运动;(c)二维纤毛搏动的动力冲程和恢复冲程(分不同阶段的正视图);(d)三维纤毛搏动的示意图和充气驱动策略;(e)其截面运动沿驱动器形成具有方向不对称的平面外运动;(f)三维纤毛搏动的动力冲程和恢复冲程的不同阶段(俯视图和正视图)。
《3.2 延展的对称破坏纤毛搏动模式》
3.2 延展的对称破坏纤毛搏动模式
就像9 + 2生物结构可以进化出丰富的运动模式一样,受内部驱动机制启发的管状驱动器也可以衍生出许多未报道的驱动策略。如图5(a)所示,原本的二维纤毛搏动是由两个交替驱动的通道延展而来的,而原本的三维搏动是通过三个通道的顺序驱动实现的。在此基础上,我们推断变化驱动器的三个通道的充气模式会产生新的搏动样式,因为这些变化会给系统带来新的非对称性。为了证明这种策略在扩展新型纤毛搏动样式方面的有效性,我们任选一个通道(通道III)作为演示,初始状态下通入气体体积设为零,在整个运动周期内设置恒定充气量、从0到更高的充气量峰值(相比于λ)和从0到更低的充气量峰值(相比于λ),并分别将它们标记为A型、B型和C型。
《图5》
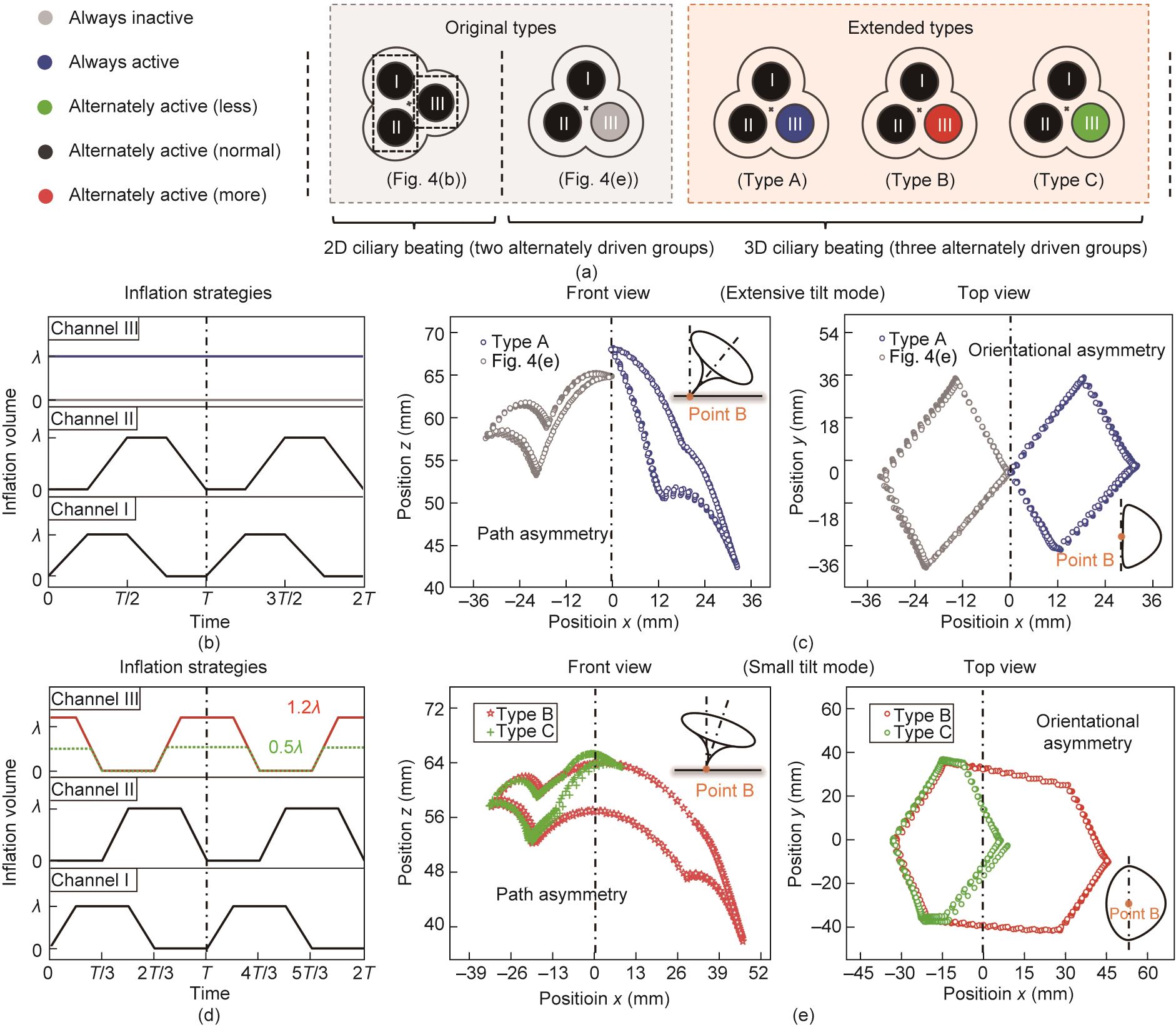
图5 基于所提出工程平台而拓展的仿生纤毛搏动策略。(a)提出的驱动策略:原本类型和扩展类型;(b)原本型和A型三维纤毛搏动的充气策略:通道I和II交替充气(充气峰值:λ),通道III充气量恒定(0或λ);(c)驱动器通过大倾斜模式实现路径不对称(正视图)和方向不对称(在俯视图中的偏心轨迹);(d)B型和C型三维纤毛搏动的充气策略:三个通道交替膨胀(通道I和II的充气峰值:λ),通道III的充气峰值调整为更大的1.2λ和较小的0.5λ;(e)驱动器以小倾斜模式实现路径不对称(正视图)和方向不对称(在俯视图中的偏心轨迹)。
对于A型搏动,我们将驱动器的通道III从零体积更改为整个运动周期中充入恒定的λ体积的气体(蓝线),同时保持其他参数不变(即通道I/II交替地充气/放气,充气体积范围:0~λ),见图5(b)。新的驱动方式破坏了各方向弯曲变形的一致性,进而导致了路径不对称(正视图)和方向不对称(俯视图),扩展了一种新型的三维纤毛搏动模式[见图5(c)和附录A中的视频S2]。对于B型和C型搏动,我们改变通道III的峰值膨胀体积(B型:1.2λ,红线;C型:0.5λ,绿线),同时保持其他两个通道的峰值体积为λ,并交替对三个通道充气或放气,如图5(d)所示。同样,充气体积峰值的变化也会引发不对称性并呈现新型的纤毛搏动模式,其兼具方向不对称(俯视图中的偏心轨迹)和路径不对称(正视图)特征,如图5(e)和附录A中的视频S2所示。
在A型纤毛搏动中,尖端轨迹仅向中心轴的一侧倾斜,底部中心[图5(c)中的B点]位于轨迹的边缘上。这种特征类似于原本的三维纤毛搏动,表现为大倾斜模式[17]。然而,由于通道III中的恒定充气体积差异(A型为λ,原本型为0),它们的尖端轨迹偏向中心轴的相反侧,且相比之下,A型纤毛搏动包络更大的区域,见图5(c)。另一方面,B型和C型纤毛搏动则表现出不同的特征。它们的尖端轨迹分布在中心轴的两侧,而非一侧,底部中心的B点被轨迹包络[图5(e)],被称为小倾斜模式[17]。此外,由于充气体积峰值的不同(B型为1.2λ,C型为0.5λ),B型比C型包络了更大的区域[图5(e)中的正视图],这些结果都进一步丰富了运动模式的多样性。值得注意的是,提出的三种新型搏动类型只是为证明所提出扩展策略有效性的部分例子。实际上,更多的变化可以被引入三通道管状驱动器来创建多样的搏动类型,如调节充气速度和三个通道的多样充气周期等。这表明引入不对称特征是为广泛的基于响应材料人工纤毛系统推导出新驱动策略的通用方法,这也与当前对生物纤毛的物理理解[18]和一些新型的仿生设计概念十分一致[20‒21,31,45]。
《3.3 低雷诺数下多种推进模式的演示》
3.3 低雷诺数下多种推进模式的演示
为了直观地展示上述鞭毛/纤毛运动的有效性,我们搭建了低雷诺数流体环境来进行一系列推进测试。这里管状驱动器被浸入硅油中,并按照上述驱动策略向三个通道中充气来执行驱动。接下来,记录红色的标记流体(在驱动器下方尖端)的运动以评估推进效率。这些实验中的预估雷诺数为Re = lf/ν = 0.014,其中,l是驱动器的长度(为65 mm),f是驱动器的驱动频率(为1/30 Hz),ν是硅油的动力学黏度(为10 000 cSt, 1 cSt = 1 mm2·s-1)。有关实验设置的更多信息详见附录A的补充文本。
记录的五种纤毛搏动下的流体推进结果如图6所示,包括初始位置(红色)、中间过程(灰色)和最终位置(蓝色)。简而言之,它们都可以在低雷诺数下有效地推进流体。具体来说,在二维纤毛搏动中,通过动力冲程和恢复冲程的不同路径形成扫掠区域,在十二个循环周期后,导致红色标记区域向左移动约8.5 mm [图6(a)和附录A中的视频S3]。对于三维纤毛搏动,有效推进是通过方向不对称实现的,这将导致轨迹偏心方向的净位移。如图6(b)~(e)和附录A中的视频S3所示,经过十二个循环周期后,原本型和A-C型三维纤毛搏动分别推动标记流体移动8.9 mm、10.1 mm、17.0 mm和14.6 mm。原本型和C型三维纤毛搏动与其他两种搏动类型相比,都分别表现出了较差的推进效果,这也与之前记录的尖端轨迹差异特征一致。此外,原本的三维纤毛搏动和C型纤毛搏动显示了推进和混合双重效果[图6(c)、(e)],这可能是由于此处标记的流体处于流体推进区域和流体混合区域的共同作用地带(详见附录A中的图S7和补充文本)。上述结果验证了仿生纤毛搏动在低雷诺数环境下推进的有效性。然而也值得注意的是,不同于非对称的纤毛搏动,因为鞭毛对称往复的摆动(对应平面波运动)和旋转(对应螺旋波运动)受斯托克斯流体动力学的物理限制,其截面运动不能直接推进流体(附录A中的图S8和S9及补充文本和视频S3)。为了实现有效推进,鞭毛需要沿细长的尾部(身体)传递截面运动以形成手性波推进。
《图6》

图6 驱动器的低雷诺数推进测试。(a)原本的二维纤毛搏动的流体推进测试实验;(b)原本的三维纤毛搏动的流体推进测试实验;(c)~(e)A型、B型和C型三维纤毛搏动的流体推进测试实验。以上实验中,搏动模式的主要运动特征和产生的推进方向见左侧的示意图。同时,这些实验演示以三个循环为间隔记录了十二个循环周期。初始位置、中间过程和最终位置分别用红色、灰色和蓝色标记。
《3.4 讨论》
3.4 讨论
理解相同的生物结构如何演化出多种运动方式的机制,为设计人工鞭毛/纤毛系统提供了灵感。如表1所示,同之前的研究[7,19‒25,27,33‒34,41‒42,44‒46]相比,本文提出的、受生物内在机制启发而制作的驱动器实现了对主要鞭毛和纤毛运动的前所未有的整合。在机制层面,我们提供了通用的设计和驱动策略来扩展现有的仿生机制[13,18,52]。尽管提出的仿生驱动器仍然不能完全复刻精密的生物内部结构,但它的确帮助我们直观地理解鞭毛和纤毛内部的运动,并可能为探索具有9 + 2或9 + 0结构运动纤毛的微管分组策略带来灵感。在仿生驱动器层面,提出的驱动器具有显著的独立寻址能力(类似于之前提出的几个纤毛启发驱动器[30‒31,45]);此外,当扩展到多个驱动器时,所具有的丰富运动使其成为极好的、可用于理论研究和验证的工程平台,来帮助我们理解生物系统中涌现的现象和原理(如多运动模式交互的流体动力学)。总而言之,所提出的一体式驱动器加深了我们对鞭毛和纤毛推进机制的理解,丰富了仿生系统的设计,并为探索生物系统的物理机制提供了一种可行的方法。
《表1》
表1 与当前受鞭毛/纤毛启发人工系统的比较
| Source of inspiration | Actuation principle | Individually addressing | Achieved flagella/cilia-like motions | Function performed | Ref. | |||
|---|---|---|---|---|---|---|---|---|
| Planar wave motion | Helical wave motion | 2D asymmetric beating | 3D asymmetric beating | |||||
| External motion | Magnetic | N/A | N/A | N/A | Yes | Yes | Propulsion | [ |
| External motion | Magnetic | N/A | N/A | N/A | Yes | N/A | Propulsion | [ |
| External motion | Magnetic | N/A | N/A | N/A | N/A | Yes | Propulsion | [ |
| External motion | Magnetic | N/A | N/A | N/A | Yes | N/A | Propulsion | [ |
| External motion | Magnetic | N/A | Yes | Yes | N/A | N/A | Swimming | [ |
| External motion | Electric | Yes | N/A | N/A | Yes | N/A | Propulsion | [ |
| External motion | Electric | Yes | N/A | N/A | Yes | N/A | Bionic | [ |
| External motion | Electric | Yes | Yes | Yes | N/A | N/A | Swimming | [ |
| External motion | Light | Yes | N/A | N/A | Yes | N/A | Bionic | [ |
| External motion | Magnetic | N/A | N/A | N/A | N/A | Yes | Propulsion | [ |
| External motion | Magnetic | N/A | N/A | N/A | Yes | Yes | Propulsion | [ |
| External motion | Magnetic | N/A | Yes | N/A | N/A | N/A | Swimming | [ |
| External motion | Magnetic | N/A | N/A | Yes | N/A | N/A | Swimming | [ |
| External motion | Magnetic | N/A | N/A | N/A | N/A | Yes | Propulsion | [ |
| External motion | Hydraulic | Yes | N/A | N/A | Yes | N/A | Propulsion | [ |
| External motion | Magnetic | N/A | N/A | N/A | Yes | Yes | Propulsion | [ |
| Internal mechanism | Pneumatic | Yes | Yes | Yes | Yes | Yes | Propulsion | This work |
《4、 总结》
4、 总结
作为一个极好的切入点,鞭毛和纤毛共有的9 + 2结构为我们提供了在一个仿生设计中模拟多种鞭毛和纤毛运动模式的机会。通过研究多种鞭毛和纤毛运动的内在驱动机制,我们提出了统一的物理模型,并进一步简化了复杂的生物结构和内部驱动策略。我们得出结论,各类鞭毛和纤毛运动可被视为由三个功能区域通过不同的驱动策略而实现。受此启发,我们制造了一体式的三通道管状驱动器以复现鞭毛的平面/螺旋波运动和纤毛的二维/三维不对称搏动。基于所提出的工程平台,我们扩展了更多的驱动策略来模拟三维纤毛搏动。最终,通过低雷诺数推进测试验证了所提出的仿生运动的有效性。综上所述,本研究为仿生鞭毛和纤毛系统的开发提供了新的设计理念和通用的驱动策略,并探索了受内在驱动机制启发的仿生学。











 京公网安备 11010502051620号
京公网安备 11010502051620号




