

《1、 引言》
1、 引言
由于全球范围内肾衰竭发病率升高的速度远超治疗方法研发的进展速度,因而催生了对有效的组织工程解决方案的迫切需求[1]。目前对肾脏功能衰竭的唯一有效治疗方法是供体移植,而唯一可行的治疗方法是透析。肾脏移植本身就依赖于供体肾脏的供应,而供体肾脏的供应在全球范围内均处于短缺状态。例如,在美国,患者接受肾脏移植的等待时间平均为3~5年,很多患者尚未等到器官移植就已离世[2]。还有,即便接受肾脏移植后恢复了肾脏的基本功能,但还需要进行免疫抑制,使患者终身都处于感染的风险中,同时还伴有不断增加的患癌风险[3‒9]。在有资格接受移植前,患者需要一直接受透析治疗,而这种治疗不能真正替代肾脏功能。同时,透析还给患者造成重大的风险,平均每8周中每100名透析患者中就有1名因透析而死亡[10]。估计到2030年,大约70%的肾衰竭病例将发生在缺乏用于治疗这种疾病的基础设施的国家[1]。因此,开发一种替代治疗方式显然是必要的。
通过肾脏组织工程方法有望开发出永久性的解决方案,解决目前治疗中存在的许多不足之处。如果能成功设计出真正的肾脏组织替代物,组织工程疗法将消除对活供体组织的依赖,从而减少患者需要透析的时间。此外,将自体原代或干细胞来源的肾脏细胞用于组织工程,可以消除对免疫抑制的需求。虽然近十年在这方面已经取得了很多进展,但是肾脏组织工程仍有许多技术和科学难题需要解决。
当前组织工程方法进展中的一个主要障碍——也是本综述的重点——是如何对已移植的组织构建体进行血管化。在任何组织工程系统中,一个成功的关键因素就是为所取得的组织提供适当的营养素、进行代谢物交换,以及氧化处理。众所周知,细胞需要在营养素和氧气供应的100~200 μm范围内才能存活[11]。在体内,已经可以通过遍布整个组织结构的微脉管系统达到这种营养素和代谢物交换。尽管在这一领域已取得了最新的进展,但是工程技术也仅限于设计复杂而精确的血管网络,以保持这种能力;在较大的组织构建体(比如供体组织替代材料)的开发设计上仍然存在障碍。本文的重点是血管化方面的问题,尤其关注组织工程中肾脏脉管系统的总体设计和整合中的最新进展,以及这些方法存在的缺点。
《2、 确定肾脏组织工程面临的血管化方面的挑战》
2、 确定肾脏组织工程面临的血管化方面的挑战
在许多组织工程应用中,建立足够的血管形成是一个普遍的挑战,在肾脏组织工程中这种挑战尤其明显,因为除了在营养素和代谢物交换方面的作用外,足够的血管形成对肾脏组织的功能至关重要。更具体地说,血液流经肾脏,通过其功能单位,即肾单位,对于肾小球过滤、血压调节、代谢物和化学平衡是必不可少的。在肾脏功能衰竭的许多病理过程中,如果没有这种血管系统的适当流动,肾脏就无法正确地清除血液中的废物,去除细胞产生的酸性物质,调节水和其他液体的水平,制造有助于产生红细胞的激素,并平衡血液矿物质水平。这种器官功能的丧失对整个身体功能和体内平衡有直接影响。例如,酸性物质和废物在体内堆积,肾衰竭导致身体失衡;红细胞生成受阻,发生贫血;血压不规律和浮肿,伴有液体水平失衡;矿物质失衡,引起骨质疏松症。肾脏血管系统需要提供的额外功能增加了该器官中血管网络的复杂性,并升级了肾脏组织工程面临的血管方面的挑战。本节关于肾脏血管网络的复杂性和整合性的概述突出了设计这样一个网络需要面临的挑战。
血液通过每个肾脏从肾门流向肾皮质和肾单位,具体过程如下:肾门处的肾动脉分支为节段动脉、叶间动脉、弓状动脉和进入肾小球毛细血管网的输入小动脉。在此处血液被过滤为初始尿产物,然后进入肾小球外的输出小动脉,进入肾小管周围毛细血管。然后这些毛细血管连接小叶间静脉和弓状静脉,进入肾静脉,将血液从肾脏输出(图1)[12]。这种脉管系统对肾脏功能有几个特别重要的方面,是设计工程组织时要考虑的关键因素。这些方面包括:①支持肾单位的肾小管周围毛细血管,同时在尿液浓缩中起着关键作用;②肾小球内对肾脏功能至关重要的三层过滤;③肾脏脉管系统与不同身体系统的整合;④肾脏血管网络必须承受的高血流量和压力。这些方面中的任何一个对于肾脏组织工程来说都会产生独有的血管化挑战,在下面进行简要的说明。
《图1》

图1 肾脏血管构造。血流通过肾门处的肾动脉进入肾脏,肾动脉分支为节段动脉、叶间动脉、弓状动脉和进入肾小球毛细血管网的输入小动脉。经过输入小动脉进入肾小球的血液以一个三层滤网过滤(包括有孔肾小球毛细血管、基底膜和足细胞足突之间的缝隙),以产生初级尿产物。血液中未过滤的部分通过输出小动脉流出肾小球,流入管周毛细血管。然后这些毛细血管连接到小叶间静脉和弓状静脉,进入肾静脉。图片由BioRender软件创建。
(1)管周毛细血管支持大部分氧气和代谢物的交换,以维持肾单位的细胞,同时在最初的尿液产物通过亨氏(Henle)环时,对液体和电解质的交换也起着至关重要的作用。如果没有这种强大的微脉管系统,肾单位的生存能力将受到影响,从功能上来说,身体将无法正常排泄废物以维持体内平衡。
(2)在肾小球毛细血管和收集最初尿产物的鲍曼氏囊(Bowman´s capsule,肾小球囊)之间高度的整合连接处存在三层过滤。该连接由肾小球的有孔毛细血管、带电基底膜和足细胞足突间的缝隙组成。系统的每一层都起到筛子的作用,留下血液中带电和较大的蛋白质及细胞,同时允许废物和水通过。研究还发现,这种三层系统的流动对于涉及肾单位组织结构的正常发育和维持的旁分泌信号至关重要[13]。
(3)由于肾脏脉管系统与鲍曼氏囊连接,脉管系统也必须与各种细胞群体适当连接,以实现适当的肾脏功能。最值得注意的是,在释放肾素的肾球旁细胞附近的肾小球门适当布置输入小动脉,对调节血压至关重要。脉管系统还与负责红细胞生成素释放和维生素D正常代谢的细胞群相连接。
(4)肾脏的所有血管组件需要承受大量的流量和压力,因为两个肾脏总共接收20%~25%的心输出量。一般来讲,经过发育的脉管系统的流动对于血管的正常发育和维持是必不可少的。
《3、 工程应用》
3、 工程应用
目前已采用许多不同的组织工程方法来解决血管网络工程的问题。在本文中,根据预期的目的,即工程复杂性和精确的细胞放置或将血管系统整合到肾脏组织结构,将这些方法分为几类。虽然其中一些方法尚未被直接应用于肾脏组织工程,并在很大程度上分散了组织血管形成的挑战,但将这些离散的方法结合在一起,可能在不久的将来会提供一种适合于肾脏组织整合的解决方案。
《3.1 工程的复杂性和精确的细胞布置》
3.1 工程的复杂性和精确的细胞布置
引入许多不同的组织工程方法来构建血管网络,包括气体发泡、溶剂浇注、颗粒浸出、纤维黏结、相分离、静电纺丝和自组装[14‒23]。虽然应用这些方法已经取得了一些成功,但仍然难以构建长期畅通的微血管网络,因为这些技术目前尚欠缺重现微体系结构所需的控制和精度。迄今为止,可成功制作的人造血管移植体仅限于直径大于8 mm的移植体。由于缺少健康的内皮细胞层、直径不匹配、合成材料表面的特性以及与天然组织的顺应性不匹配,导致内膜增生、血栓形成或移植体感染[24],较小的血管替代物(< 6 mm)通常无法保持长期畅通和生存能力。有研究者认为,通过更好地控制支架微结构和细胞布置,可以克服其中一些问题。
由于这些原因,最近的研究重点是开发生物打印技术,使用生物相容的载细胞生物墨水,取得更高水平的制造控制。图2显示了最常见生物打印方法的示意图。有必要对支架结构的构建和细胞布置采取更高水平的控制,以重现肾血管网络的复杂结构。最近很多研究深入探讨了当前的血管生物打印方法[24‒26]。本文简要概述了主要的生物打印策略,旨在为组织工程制造微血管,包括基于挤压的生物打印、基于液滴的生物打印和基于激光的生物打印,所有这些策略的主要区别在于它们提供的控制级别。
《图2》
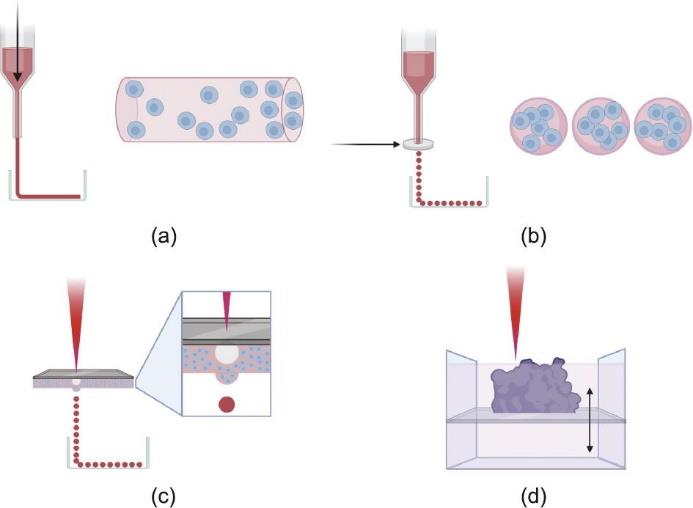
图2 主要生物打印技术概述。(a)挤出式生物打印:采用气动、活塞或螺旋机构向注射器中的生物墨水施加压力,在平面上打印出各种形状。同轴或多轴喷嘴可用于挤出中空形状,以及具有更复杂结构的形状。(b)液滴式生物打印:采用热、静电、压电或微阀技术(图中显示的银色圆盘),释放载有细胞的水凝胶液滴。图中(a)和(b)部分右侧显示了液滴式技术相对于挤出方法在均匀的细胞和支架分布上的优势。(c)激光式液滴生物打印:采用钛或金薄膜和激光,在生物墨水底层产生气泡,将生物墨水液滴推到表面上。(a)~(c)部分所示的方法可用于支撑槽中,以获得更好的稳定性,可用于挤压随后用于间接生物打印方法的牺牲材料。(d)激光式光固化技术:在生物墨水槽中设计一个多孔平台,利用激光固化来逐层构建结构。图片由BioRender软件创建。
《3.1.1. 挤出式生物打印》
3.1.1. 挤出式生物打印
挤出式生物打印方法的主要原理是将含有或不含细胞的生物墨水通过注射器筒挤压到表面上,从而沉积成预编程的图案。根据材料特性,分配的生物墨水通过物理或化学交联发生凝固。为获得生物墨水布置的高精度和所得结构的完整性,生物墨水通常需要剪切稀化水凝胶,使水凝胶可以容易地从打印机的分液喷嘴挤出,并且在挤出时必须快速固化。生物墨水还需要具有生物相容性,并支持细胞的存活和生长。这些要求也限制了可使用的生物墨水的材料选择。利用挤出式喷嘴不同的尺寸和类型,还能额外增加打印结构设计的复杂性。
直接挤出式生物打印方法可将生物墨水布置成不同图案,这些图案自身将构成血管组织。这些图案可以通过以不同生物墨水和细胞的镶嵌来进行复杂的设计,取得特定细胞类型和材料的精确布置。血管生物打印的早期研究主要集中在以挤压生物墨水或搭载细胞的生物墨水形成管状形状的方法上。在这些研究中,使用巯基化透明质酸和明胶凝胶直接将NIH 3T3细胞打印成管状结构[27‒28]。这些管状结构的打印证明了使用生物打印来创建血管和其他管状结构的可行性。还有一些其他研究小组采用不同的方法创建了管状结构,其中几个研究小组试图垂直地生物打印大直径的独立管状结构[29]。尽管在打印这些形状方面取得了成功,但是这些结构不能长期存活,主要原因是结构不稳定。最近开发的具有增强结构稳定性的生物打印方法主要集中在为支架提供临时支撑,直到发生完全交联,或者提高打印过程中交联的速度和强度。例如:Hinton等[30]试图通过开发在辅助水凝胶槽中打印结构的技术,稳定生物墨水的布局,从而增强结构的完整性。这种技术可防止生物墨水在挤出后,在交联发生前就开始扩散。
很多研究小组利用改进的交联方法,使用同轴喷嘴,挤出涂覆了另一种材料的芯材料,以一个步骤将中空纤维打印到微脉管系统范围的表面上。比如:Yu等[31]开发了同轴挤出,然后瞬时交联以支撑管状纤维的方法。这些研究人员证明了利用这种生物打印中空纤维的方法进行载细胞生物打印的潜力,因为使用这种方法打印的人脐静脉平滑肌细胞在灌注培养7天后取得了84%的细胞存活率[31]。Dolati等[32]对这些方法进行了改进,以强化管状纤维的完整性。通过开发融合相邻中空纤维的方法对同轴方法进行进一步改进,使管状网络更近似于自然血管网络[33]。最近的研究扩展了同轴喷嘴的概念,以新开发的多轴挤压喷嘴在凝胶支架内打印双层和三层中空通道,创建更高仿生程度的网络[34]。器官微结构的计算机辅助设计(CAD)模型也被用于挤出式生物打印,模拟仿生血管构建体[35]。
稳定挤出血管结构的另一种方法是利用以间接生物打印方法取得的支撑支架的稳定性。间接挤出式生物打印方法是将牺牲材料(如明胶、藻酸盐、琼脂糖或普兰尼克)按所需的血管通道图案,打印到周围有或没有细胞的基质材料中。通常,牺牲材料随后会被移除或洗掉,然后用内皮和(或)血管平滑肌细胞接种中空血管树。Lee等[36‒37]最近采用这种方法制作了一个与两条较大血管吻合的微血管网络。研究人员将明胶通道打印到胶原蛋白基质中,该基质与嵌入了人脐静脉内皮细胞(HUVEC)和肺成纤维细胞的纤维蛋白床相邻。通过加热除去明胶,并用内皮细胞接种中空管。对整个结构培养数周,产生与较大血管吻合的微脉管系统。
挤出式生物打印技术从能打印大血管、微血管发展到现在能分支微血管,使人们离获得为组织工程应用综合设计和创建微脉管系统的能力又进了一步。虽然这些构建体的稳定性和长期生存能力正在不断提高,但是这项技术还存在一个重要的局限性,就是需要取得所需的控制水平。从理论上讲,生物墨水应该均匀分散在挤出的生物墨水构建体中,但是,在实际操作中,细胞和水凝胶分布的均匀性会变差。如上所述,由于高度的复杂性以及脉管系统和组织结构之间精确界面整合的必要性,这种精确控制的缺失成为了肾脏应用中的一大问题。因此,重要的是要考虑更好地控制细胞布置的方法,这涉及液滴式生物打印和激光式生物打印。
《3.1.2. 液滴式生物打印》
3.1.2. 液滴式生物打印
液滴式生物打印(直接和间接)的核心方法类似于挤出式生物打印。二者的不同之处在于,液滴式生物打印是将包含特定数量细胞的小液滴而不是稳定的材料流布置在图案中。这种方法的优点是所得的构建体在细胞分布和材料组成上更均匀。这些液滴以热、静电、压电或微阀技术形成,每次推出特定尺寸和数量的液滴[35]。在一项有关直接液滴式生物打印的早期研究中,研究者采用喷墨生物打印,对这种技术可用于制造管状水凝胶构建体进行了概念验证[38]。该研究小组的结果还表明,包裹在这些构建体中的人类造血干细胞可以在培养过程中分化成多种细胞类型。Cui等[39]进行的后续研究表明,液滴式生物打印可以与人类微血管内皮细胞结合使用,以制造具有融合血管内衬的微血管构建体,该构建体显示出强劲的细胞增殖和细胞间黏附。另一项研究表明,通过将藻酸盐液滴打印到氯化钙池中,可以更好地控制这些技术,使得钙快速扩散到藻酸盐溶液中,从而在接触时使其稳定,这与上述挤出式生物打印实例中的交联固化非常类似[40‒41]。还有其他研究采用直接液滴式生物打印,将藻酸盐液滴打印到氯化钙池中,该技术可用于生物打印更复杂的分支构建体,以便更准确地再现天然血管结构[42‒43]。这些研究小组在他们构建的构建体中取得了较高的精度和细节,以及培养后较高的细胞活力。
间接液滴式生物打印的操作方法与挤出式生物打印方法非常类似。比如:在上述挤出式打印实例中证明微血管床形成的研究小组还用液滴式方法进行了类似的研究。他们在研究中将明胶和人脐静脉内皮细胞的混合物制成液滴,沉积到与人脐静脉内皮细胞和肺成纤维细胞的纤维蛋白床相邻的两根血管中[37]。然后将整个结构加温,溶化明胶,继续培养,让人脐静脉内皮细胞附着到血管的内衬上。这样就形成了融合内皮衬里[37]。
《3.1.3. 激光式生物打印》
3.1.3. 激光式生物打印
激光式生物打印采用细胞转移技术或光固化技术来构建组织构建体。细胞转移技术的原理是通过将激光束中的能量转换为热能,使生物墨水液体物理弹射到表面。简单地讲,是将激光束发射到一层生物墨水顶部的金或钛层上。金或钛层在精确的接触区域吸收来自激光束的能量,并产生热量,蒸发少量的生物墨水,从而形成高压气泡,而该气泡迅速将气泡下方的生物墨水液滴推到表面上,在接触时进行交联[35]。虽然这种液滴形成机制比上面讨论的液滴式生物打印方法更加精确可控,但是液滴在最终图案化以产生三维(3D)构建体时也采用了相同的概念。
光固化技术采用一池或一层光敏生物墨水,生物墨水在受到照射时会固化[24]。以特定模式发射激光束,固化生物墨水的隔离区域,随后移除未固化的墨水,仅留下固化的3D构建体。如果要制作更厚的组织构建体,可以使用多孔台,使高能激光束不会消散,而是用于一层一层地固化生物墨水并将生物墨水堆积起来。尽管这种技术在提供更可控、更精确的生物打印血管构建体方法方面尚有巨大潜力,但是将该技术用于血管生物制造的研究并未取得成功,开发出的结构在完整性和细胞生存力方面未达到上述其他生物打印方法的水平。
这些生物打印方法的不同特性比较见表1。尽管这些生物打印方法已被成功用于创建简单的血管网络,但它们在肾脏组织工程中的应用仍然存在结构分辨率和完整性的问题,而这些结构分辨率和完整性是重现构成肾脏肾小球毛细血管和肾小管周围毛细血管(直径5~10 µm)的关键血管结构所必需的[44]。具体地讲,肾小球的三层滤网由有孔毛细血管(孔直径70~100 nm)、带独特电荷的基底膜(厚240~370 nm)和有孔足细胞足突(宽25~60 nm)组成,而所有这些都超出了当前打印分辨率水平[45‒51]。到目前为止,已经开发出了使用新型生物墨水再现具有某种复杂程度的肾小管结构的3D生物打印方法,可取得更好的结构完整性以及肾环境生物学相关性[17,52‒53]。在一些研究中还打印了模拟近端管状组织和几何形状的可灌注结构[53]。但是,肾小球的复杂性仍然超出当前的研究能力水平,还没有哪个研究成果能够再现肾脏脉管系统从而在一定程度上实现真正的过滤功能。另外,打印的血管网络的完整性不足,难以在流动状态下成功培养这些网络,而这一过程对脉管系统的适当成熟至关重要。重要的是,仅使用生物打印方法将血管结构人工整合到肾脏结构中仍然存在困难——这一点对于实现适当的组织存活和过滤功能至关重要。
《表1》
表1 挤出式、液滴式和激光式生物打印方法的比较
| Properties | Extrusion-based bioprinting | Droplet-based bioprinting | Laser-based bioprinting |
|---|---|---|---|
| Printing speed | 10‒50 µm·s‒1 | 1‒10 000 droplets·s‒1 | 200‒1600 mm·s‒1 |
| Resolution | 100 µm‒1 mm | 50‒300 µm | 50 µm |
| Accuracy | Medium to low | Medium | High |
| Structural integrity | High | Low | Low |
| Scalability | Yes | Yes | Limited |
| Cost | Medium | Low | High |
《3.2 脉管系统与肾脏组织结构的整合》
3.2 脉管系统与肾脏组织结构的整合
如上所述,目前通过合成创建血管网络的方法还不能按所需的分辨率再现肾脏的天然血管网络。此外,这些方法不支持血管网络与必需的肾脏组织细胞群的整合。由于存在这些局限性,最近出现了一股研究热潮,重点是以天然肾脏组织的微观和宏观结构(称作细胞外基质,ECM)作为肾脏组织工程的基础。细胞外基质是组织的无细胞支架,根据生化信号[如生长因子和血管内皮生长因子(VEGF)]提供物理组织结构和局部细胞微环境,以取得适当的细胞维持、发育和血管内向生长。本节讨论了在这种肾脏组织血管化方法方面取得的进展,下面讨论的相关基本技术的示意图见图3。
《图3》
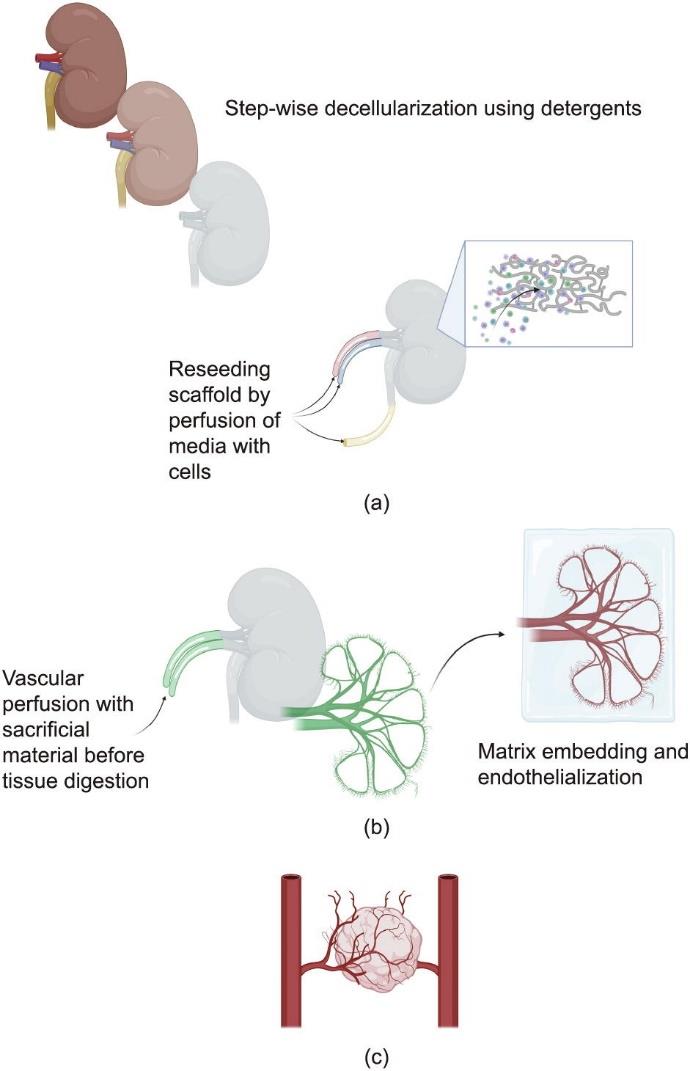
图3 使用天然组织结构进行血管化的方法。(a)脱细胞和再接种方法:首先用去污剂进行一系列清洗,使肾脏脱细胞,然后用不同细胞类型对这些支架进行再接种。这种方法依靠残余细胞微环境,将细胞引导到合适的组织位置。(b)血管铸型法:使用脱细胞肾脏内的血管网络灌注牺牲材料,在消化掉其余组织的同时,保留牺牲材料。然后将这种牺牲材料涂覆或布置在载细胞或未载细胞的基质材料中;接下来,将其移除,留下天然肾血管网络的中空铸型。(c)器官样血管化方法:利用器官样的天然VEGF-VEGF受体信号,从体内或体外的周围区域调用脉管系统。图片由BioRender软件创建。
《3.2.1. 全器官脱细胞和再细胞化》
3.2.1. 全器官脱细胞和再细胞化
在使用全器官支架作为肾脏组织血管化的基础时,中心范式是从完整的器官或组织中去除细胞碎片(称作“脱细胞”过程),随后将细胞重新接种到支架上(称作“再细胞化”过程)。从器官中去除细胞碎片时,会去除许多免疫原性反应,因为免疫反应性供体DNA和人类白细胞抗原将被去除。此外,细胞外基质材料完全具有生物相容性[54‒57]。这些脱细胞肾脏的血管网络框架必须是仿生的,并具有肾脏组织所需的分辨率。从理论上讲,细胞微环境中提供的先天细胞信号也能进行不同细胞群的适当定位,以便与肾单位的不同区域进行充分的血管整合。下面,将详细介绍已开发出的可成功实现肾脏脱细胞和再细胞化的方法。
Crapo等[58]建立了一套充分去除细胞的标准,这是开发成功的脱细胞方法的关键;他们建议使用4′,6-二脒基-2-苯基吲哚(DAPI)和苏木精-伊红(H&E)染色,同时结合凝胶电泳法,目测评估细胞的去除情况,对脱细胞组织中的DNA含量进行量化。他们的研究结果表明,如果获得每毫克组织干重50 ng的双链DNA,即表明已取得充分的脱细胞[58]。另外,少量残留细胞的支架中的任何残留DNA的长度应小于200个碱基对[58]。达到这些参数要求,即可降低支架的免疫原性,并使支架获得进一步应用。
对于支架的再接种,已经发现,保持细胞外基质中胶原、层黏连蛋白和生长因子的完整性至关重要。细胞外基质成分得到保存后,迁移、维持和适当的分化所需的细胞微环境也得到保持。以前认为细胞外基质的生长因子和趋化因子在接种细胞的结局中起着最重要的作用。但是,在最近关于细胞外基质的生物物理特性(如其刚度和固有的压电特性)与细胞结局之间关系的研究中,突出显示了保持细胞外基质结构完整性的重要性[59‒63]。由于这些原因,将有关脱细胞的方案重点放在从支架中去除足够量的细胞碎片,同时保持关键的细胞外基质蛋白和结构特性。
要在脱细胞、去除细胞碎片和保存细胞外基质结构这些目标中取得平衡是有难度的,因为如果采用快速、成功地从器官中去除细胞碎片的方法,通常会损害细胞外基质;而如果采用主要目标是保持细胞外基质结构的方法,则通常不能去除足够量的细胞碎片。例如:主要使用Triton X-100进行支架脱细胞的肾脏脱细胞方法,可成功地保持细胞外基质的生长因子和结构完整性,但不能充分去除细胞。主要使用十二烷基硫酸钠(SDS)的脱细胞方法已证明能够充分去除细胞碎片,同时保持细胞外基质的蛋白质和结构[54‒55,57,64‒66]。目前,许多有效的脱细胞方法使用Triton X-100和十二烷基硫酸钠的混合物,以取得细胞清除和细胞外基质维持的最佳水平。Orlando等[57]通过扫描电子显微镜(SEM)成像,证明了这种脱细胞方法保持了肾单位的肾小球结构,这对肾脏的过滤功能至关重要。值得注意的是,根据相关报道,这些清除方法会降低肾脏细胞外基质的刚度,并影响重新接种的干细胞的细胞结局。
一旦肾脏成功脱细胞,马上要面临的是重新接种血管网络的挑战。本文主要讨论最近有关证明脱细胞肾脏支架血管成分成功再接种的研究。再接种肾血管细胞外基质最常见的方法是通过肾动脉顺行灌注。很多研究团队使用灌注泵实现了灌注。由于血管树复杂而精细,尤其是在肾小球水平,因此确定最佳灌注压力和流速对再内皮化的成功至关重要[67]。不完全的再内皮化会带来体内血栓形成的显著风险,损害血管网络的完整性,并因此损害工程组织的长期生存能力。主要使用基于灌注的方法来重建肾脏脉管系统和肾脏组织是全器官肾脏组织工程最成功的尝试之一。在另一项研究中,进行了大鼠肾脏的脱细胞:使用肝素化磷酸盐缓冲盐水(PBS),以30 mmHg(1 mmHg = 133 Pa)动脉压力顺行动脉灌注15 min,清除肾脏的残余血液;然后以30 mmHg的恒定压力,以加入去离子水的1%十二烷基硫酸钠灌注12 h,单独用离子水灌注15 min,然后以加入去离子水的1% Triton X-100灌注30 min [65]。接下来,将悬浮在培养基中的人脐静脉内皮细胞以1 mL·min‒1的恒定流速通过动脉,接种到支架中[65]。让细胞附着过夜,之后恢复灌注培养。血管灌注后,将大鼠新生肾脏细胞通过输尿管接种到支架中,同时在器官室中的支架上施加压力梯度,使支架更好地渗透到肾脏中。然后将整个肾脏置于定制的生物反应器中,通过肾动脉以1.5 mL·min‒1的速度进行持续灌注[65]。让输尿管和肾静脉被动引流。所得组织的成像显示,细胞在被定位到正确的细胞微环境中时取得了不同的成功结果。这里值得注意的是,内皮细胞已正确定位,成功地对构建体的血管进行了再内皮化,且形成完整的细胞层。这些工程构建体随后的原位移植显示,在与肾动脉和静脉吻合后,该结构成功灌注,短期内没有血栓形成或出血。其他研究小组已经使用不同类型的细胞对肾脏脉管系统进行再接种,取得了不同程度的成功。他们还将小鼠胚胎干细胞(ESC)接种到脱细胞大鼠肾脏的肾动脉中,并在没有任何额外生长因子的情况下培养。这些细胞成熟为光滑的薄片,在血管结构中具有正确的形态[68‒69]。另有一个研究小组也将小鼠ESC接种到大鼠肾脏支架的肾动脉中,以评估肾小球和肾小管周围毛细血管的再增殖。他们发现,这些细胞最早在接种24 h后,就表达Tie-2和分化簇31(CD31)内皮细胞标记物,而且CD31表达在72 h后增加[70]。诱导多能干细胞(iPSC)衍生的内皮细胞也被用作肾血管再接种的细胞来源。在这些研究中,iPSC衍生细胞成功在血管结构内形成一层细胞层,更重要的是,也在肾小球结构内形成一层细胞层[66,71]。
然而,对于所有这些血管再植入方法,长期血栓形成的发生仍然是一个重大挑战。几个小组在灌注时应用了抗凝剂,但收效甚微[57,72‒73]。最近,一个研究小组将肝素固定到脱细胞肾脏支架的细胞外基质上,观察到与非肝素化对照和灌注时注射抗凝剂相比有改善的结果[74]。这种结果无法长期保持;尽管如此,他们的技术可能会在未来得到进一步发展以获得更好的结果。此外,仅由细胞壁龛提供的定位信号似乎不足以促使肾脏细胞正确填充支架,这可能是由于细胞无法自由移动通过这些支架。这种不足阻碍了将肾脏细胞群与血管网络正确整合的能力,尽管这种方法产生的血管网络本身比使用上面列出的生物打印方法产生的分辨率更高。
《3.2.2. 血管铸型》
3.2.2. 血管铸型
最近开发了一种新型的替代方法,将天然血管结构用于组织工程用途。这种方法是在血管铸型方法中发现的,旨在解决当前方法中存在的问题,即缺乏生物打印方法的分辨率、不成功的长期通畅率,以及与肾单位结构更好整合的能力。在该研究中,Huling等[75]旨在构建一种仿生血管支架,可以通过与整个肾脏细胞外基质进行隔离来构建这种支架。这种方法的好处是可以成功在肾小球水平上构建血管树,并且在应用于不同的水凝胶或支架以形成组织之前,可以让该血管树更完全地成熟。在预先发育的血管床周围构建肾脏组织,就有可能让更大的组织构建体在体外发育,并且当构建体在体内吻合时,有望得到更好的整合。
血管树的制作方法如下:用1 mL丙酮灌注成年大鼠的肾动脉,然后立即用1 mL溶于丙酮的10%(浓度)聚己内酯(PCL)灌注[75]。让丙酮在室温下蒸发24 h,然后在4 °C下蒸发三天[75]。蒸发后形成天然血管树的固体血管铸型。随后,将肾脏放入20%(浓度)氢氧化钠溶液中48 h,去除血管铸型周围的组织,然后用去离子水清洗24 h [75]。从现已分离的血管树腐蚀铸型中,在叶动脉水平分离片段,用9.8 mg·mL‒1的1型鼠尾胶原浸涂,然后在干燥后置于10 mmol·L‒1的1-乙基-3-[3-二甲基氨基丙基]碳化二亚胺盐酸盐和10 mmol·L‒1 N-羟基琥珀酰亚胺的交联溶液中30 min [75]。然后以温热的丙酮将PCL铸型从结构中溶解出来,留下相对容易操作的中空胶原基血管支架。简单地涂覆悬浮在培养基中的支架表面,将由绿色荧光蛋白转染的MS1内皮细胞接种到支架上。由此产生的仿生树为一种3D分支结构,具有相互连接和连续的可见中空通道。血管支架顶端的SEM图像显示肾小球结构似乎是完整的。然后将由这种内皮细胞包被的血管支架放入与肾脏细胞混合的胶原水凝胶中。在进一步的体内研究中,在外科手术造成皮质缺损后,将整个构建体移植到大鼠肾脏上。
尽管前景看好,但是这项研究仍处于早期阶段。体内研究的结果证明了这种新方法的潜力,因为在研究中发现移植的肾脏中血管生成显著增加,更重要的是,显示了血管构建体中红细胞和有核细胞的存在——这表明支架和宿主脉管系统之间发生了一些吻合。在今后有关血管铸型方法的研究中,仍然需要解决重新接种和支架成熟方法的优化问题。研究中发现,在植入后第14天,大多数MS1细胞已经移出支架,进入周围组织,表明细胞黏附于支架的不稳定性[75]。随着内皮细胞离开支架,会造成显著的血栓形成风险,因为下面的基底膜可能暴露出来,开始凝血级联反应。有研究者提出,通过血管支架内部接种内皮细胞可能比外部接种产生更好的结果,尽管灌注可能会损伤脆弱的血管结构并导致血栓形成(这种情况也发生在全器官再细胞化方法中)。另一个可能的改进是在将支架放入组织水凝胶之前,让血管支架进一步成熟,形成更强的连接,但是,血管网络的通畅性在这里可能是一个问题。
《3.2.3. 类器官血管化》
3.2.3. 类器官血管化
还有一种更好地整合工程血管网络和肾脏组织构建体的方法,即通过提升对发育中的血管形成和肾脏组织器官发生的理解,让这两种成分在胚胎发生阶段的发育期间一起生长。在发育过程中,肾脏的血管网络是通过血管发生和血管生成的结合而形成的。血管发生是从源自发育组织本身的中胚层细胞起源的血管细胞形成原始脉管系统,而血管生成是从围绕发育组织的已有脉管系统重塑和向内生长血管。有关肾脏发育的几项研究表明,脉管系统的血管生成源比血管发生源具有更决定性的影响。但是,不能忽视第二种细胞群的重要性,因为这些细胞群可能产生更强的血管向内生长的趋化信号[76‒81]。利用这两种机制的混合,可以创建一个整合到早期肾单位结构的血管网络。这是因为早期肾单位结构的发育产生了一个称为血管裂隙的区域,其中早期足细胞样细胞参与VEGF-VEGF受体信号传导,从而产生发育中的内皮细胞群[82‒84]。一旦整合并成熟,内皮细胞将成为有孔毛细血管,是肾小球和鲍曼氏囊之间三层过滤单元的一部分。
可参与VEGF-VEGF受体信号传导的发育中的肾单位结构,可以以肾器官样的形式在体外培育。类器官是产生种子组织所必需的细胞类型的集合体。不同的细胞组合可制作具有不同功能的组织。比如:已经研制了只含分离结构的肾脏类器官(比如肾小管),也设计了其他含多种肾脏细胞类型的类器官。根据这种情况,本研究将重点放在模拟胎儿肾脏组织的类器官上。该类器官由能够构建肾小管和肾小球结构的祖细胞群组成。
如上所述,血管生成在胚胎发生期间在发育肾脏的血管化中起着主要作用。因此,有研究者认为如果在体外成功构建了具有血管生成潜力的血管网络,将这些类器官一起培养可能会产生可通过灌注维持的血管化组织[85‒87]。另有研究表明,根据一些制作这些胎儿肾样器官的方案,可以将器官样细胞群发育成内皮细胞,这可能引起血管生成,因为血管生成发生在胚胎形成过程中[88]。目前的情况显示,在高流量环境中刺激类器官时,这些新生内皮细胞已经发育成熟,成为相互连接的血管床[88]。但是,形成的脉管系统自身不够强健,不足以使整个组织构建体血管化,血管网络的扩张仅侵入少数肾小球结构[88]。在成功进行体外血管生成脉管系统之前,可以了解更多关于肾脏祖类器官如何连接到外部脉管系统,并移植到体内发育的信息。
《3.2.3.1. 胎儿和胚胎干细胞来源的类器官》
3.2.3.1. 胎儿和胚胎干细胞来源的类器官
在几项体内研究中,已经将由不同细胞类型组成的肾脏类器官布置在高度血管化的区域,如网膜、前房或肾包膜下[79,89‒95]。其中的一个实验是用小鼠或猪的后肾间充质(早期肾单位结构的一种前体)进行。这些移植的后肾生长并分化为成熟的肾单位样结构,有些时候产生初步的尿样产物[89‒90,92‒94]。这些研究确定了肾脏祖类器官应具备的主要特性,以便能将这些器官发育成成熟的肾脏组织。在这些研究之后,还有其他的研究团队对包含干细胞来源后肾样细胞的类器官进行了移植。干细胞来源类器官的使用有可能在将来的研究中取得增强的可扩展性以及更广泛的应用。
有一个研究团队对小鼠胚胎肾的细胞(E11.5)进行了分离,然后又进行重组,形成了类器官。将这些类器官移植到大鼠肾脏包囊下(发育)三个星期,然后收获[96]。结果发现,虽然由细胞制成的类器官与后肾非常类似,但是类器官中血管化水平非常低,肾结构成熟不完全。关于后者(发育不完全)的证据是:内皮细胞经常缺乏窗孔,以及肾小球基底膜覆盖不完全,具体见透射电子显微镜(TEM)影像。后来进行相关研究对这些结果进行优化,确定血管化和肾脏细胞成熟上的差异可能是由移植体中产生的VEGF不足造成的[96]。利用生长因子对移植体进行处理后,可显著改善血管化和肾脏细胞成熟的结果。在固定之前,将异硫氰酸荧光素(FITC)结合的牛血清白蛋白(BSA)和FITC标记的葡聚糖注射到宿主脉管系统中,然后在管状结构和肾单位中进行检测,以测试这些类器官的过滤功能。发现牛血清白蛋白和葡聚糖在近端肾小管结构中与巨蛋白共定位,表明它们已经成功地被肾小球过滤,并被小管内吞。但是,对于这些结果的另一种解释可能是,由于内皮屏障不完全成熟,牛血清白蛋白和葡聚糖只是渗漏到管状结构中。
另有一个研究组将E13.5小鼠的胚胎干细胞来源类器官与人脐静脉内皮细胞和间充质干细胞一起使用[97]。值得注意的是,研究组将这些类器官植入无胸腺小鼠的颅骨中后,发现类器官迅速血管化并成熟。注射荧光黄后,以活体显微镜观察肾小球灌注和肾小管过滤[97]。与以前的研究相比,通过这种方式可在研究中对植入的类器官进行更实时的血管化成像和功能评估。
《3.2.3.2. iPSC来源的类器官》
3.2.3.2. iPSC来源的类器官
由于使用胚胎来源细胞的伦理问题和政府法规,用于组织工程用途时,可能使用人类诱导多能干细胞(hiPSC)比胚胎干细胞(ESC)更易转化。最近在肾脏类器官移植上取得的突破大多数都是采用人类诱导多能干细胞来源的肾脏祖类器官。人类诱导多能干细胞是去分化的成体细胞,最常见的是皮肤成纤维细胞。这些去分化细胞暴露于一系列或多种因子,这些因子引导细胞向中胚系分化,并进入肾祖细胞轨道,向后肾样终点分化。已确定了几个成熟的分化方案,但都是基于对肾脏胚胎发生中的发育线索的深刻理解,并将人类诱导多能干细胞推向相同的分化路径[98‒100]。总的来说,产生的类器官在肾脏细胞群成熟度和比例方面存在差异,基于聚集技术获得的结果在可重复性和可变性方面存在一些差异。
本研究着重于体内血管化环境中这些肾祖类器官的应用。在这些类器官在体内应用的最新进展中,大多采用了肾肩胛下移植的方法。Sharmin等[101]将这些类器官与浸泡在VEGF中的两根棒状琼脂糖植入物、人脐静脉内皮细胞的聚集体和间充质干细胞一起移植到无胸腺小鼠的肾包膜下。在移植后,类器官中的肾小球被宿主脉管系统血管化,并生长成熟20天。对肾小球的内皮细胞开孔,完整的双层肾小球基底膜已经形成,并且足细胞已经形成足突,以形成上述的三层筛,这样才能保留必要的成分(如红细胞和大蛋白),同时过滤废物,从而形成基本的尿产物。在随后的研究中使用类似的方案移植了这些类器官,但没有添加外部VEGF,结果证明人类诱导多能干细胞来源的肾类器官能够产生自己的VEGF [99]。即使没有添加外部VEGF,类器官构建体也将血管化,并成熟发育为肾结构。
后来Van den Berg等[102]进行了一项研究,在小鼠的肾包膜下移植了类似的肾祖类器官,但没有任何其他支持细胞群。在该研究中,通过外科手术放置的腹部成像窗,使用活体体内成像,重点评估取得的肾脏组织的血管网络和肾单位结构的时间成熟度。他们还使用TEM影像评估了肾脏微架构的成熟度。这些研究者证明了在7~28天的观察期内,脉管系统的渐进向内生长,以及类器官的肾小球和管状结构的成熟[102]。还使用TEM影像显示三层筛的发育情况:肾小球、基底膜和足细胞足突间的缝隙。另外,管状结构的TEM影像中还显示以前没有评估过的顶端刷状缘成熟。与Sharmin等[101]使用的方法相比,使用该研究小组的方法观察到的肾脏结构成熟度有所不同,即足细胞足突在结束时间点不太成熟。
最近,Francipane等[103]按照Takasato等[98]的方案,将人类诱导多能干细胞来源的肾类器官植入淋巴结。之所以选择淋巴结作为植入部位,原因是淋巴结比肾包膜更容易进入,并且之前已经发现淋巴结支持成年小鼠肝脏、胰腺和胸腺细胞的植入和功能[104]。此外,Francipane和 Lagasse [105‒107]之前发现淋巴结环境可以支持胚胎和胎儿肾脏组织功能的成熟。移植的类器官在一周内显示出宿主内皮细胞浸润的迹象,并在移植后6周内显示出含有宿主来源的CD-31阳性内皮细胞的肾小球样结构的发育。这表明正在发生血管生成,并且脉管系统正在被很好地整合到发育中的肾单位结构中[103]。这项以淋巴结作为肾类器官移植位点的研究尚处于早期阶段,还存在包括脱靶细胞群在内的问题。但是,该研究已证明工程肾脏组织在血管化方面的巨大前景。
还有,可以从这些肾脏组织工程血管化的类器官研究中获得相关的知识,即理解工程血管网络如何被整合到肾结构中,以成功重建过滤功能和长期的组织存活力。表2 [76‒77,79,94,96,101‒103,108]中列出了肾脏类器官移植的一些主要方法,以及由此产生的脉管系统的质量。准确评估血管化类器官的过滤功能仍然存在困难,因为体内植入时,它们不能被分离。因此,开发体外血管系统将很有价值。另外,对肾实质的移植研究可能对将来评估类器官脉管系统如何被整合到肾脏脉管系统中的研究有好处。还有,目前的移植方法显示在植入区域有大量的脱靶细胞群。许多此类脱靶细胞具有软骨形成形态,表明在这些植入的构建体中可能存在低氧区域,因为中胚层干细胞在低氧条件下易于分化成软骨细胞。这一发现说明了对更快的血管形成或桥接技术的需求,在使用干细胞来源的组织工程方法进行血管整合的早期阶段,提供了替代的氧合途径。
《表2》
表2 不同类器官移植方法造成的血管贡献
| Reference | Cell source | Transplant location | Treatment duration | Origin of resulting vasculature |
|---|---|---|---|---|
| Hyink et al. [ | Mouse metanephroi E11/12 | Anterior eye chamber | 6 days | • Microvessels predominantly donor • Larger vessels predominantly host |
| Robert et al. [ | Mouse metanephroi E12 | Under kidney capsule, neonatal and adult | 6‒7 days | • Neonatal: combination • Adult: predominantly donor |
| Loughna et al. [ | Mouse metanephroi E11/12 | Neonatal nephrogenic renal cortex | 7‒10 days | • Predominantly donor |
| Rogers and Hammerman [ | Rat metanephroi E15 | Omentum | 2 weeks | • Partially host, did not stain for donor cells |
| Dekel et al. [ | Human metanephroi 7‒8 weeks | Under kidney capsule | 4 weeks | • Partially host, did not stain for donor cells |
| Xinaris et al. [ | Embryonic mouse kidney organoids E11.5 | Under kidney capsule | 3 weeks | • Donor |
| Sharmin et al. [ | hiPSC-derived kidney organoids | Under kidney capsule | 20 days | • Host |
| Francipane et al. [ | hiPSC-derived kidney organoids | Lymph node | 6 weeks | • Host |
| Van den Berg et al. [ | hESC- and hiPSC-derived kidney organoids | Under kidney capsule | 7‒28 days | • Combination |
《4、 结论和未来展望》
4、 结论和未来展望
对组织工程血管化方法的回顾表明,有大量研究旨在解决这一核心挑战。仅在过去5年中,无论是单独的还是应用于肾脏组织工程领域,在生物打印、脱细胞、再细胞化、血管铸型和类器官发育等领域取得了许多进展。生物打印方法的进步已经成功地产生了独立的更大直径的血管和微血管,现在专注于建立一个优越的微血管仿生网络。去细胞化、再细胞化、血管铸型和类器官开发的进展旨在提高生物打印方法的分辨率,重要的是,要证明血管网络如何被更好地整合到肾脏组织中。很明显,当前方法中仍有待解决的关键缺点,即生物制造方法的分辨率和保真度不高、血管组织被整合到肾脏组织的策略不足,以及工程血管网络的长期通畅性。在解决了这些缺点之后,剩下的挑战将是在高流量下长期灌注这些结构。正如本文第 2 节所述,肾脏的所有血管成分都必须承受大量的流量和压力,因为两个肾脏一起接收20%~25%的心输出量。这种流动很重要——首先,对于肾脏血管系统的正常成熟;其次,对于工程肾脏的正常功能。尽管剩下的挑战并非微不足道,但现在比以往任何时候都更容易实现。其中许多进步为未来可能用于改善全球患者生活的技术带来了希望。











 京公网安备 11010502051620号
京公网安备 11010502051620号




