
2017, Volume 3, Issue 3
Engineering >> 2017, Volume 3, Issue 3 doi: 10.1016/J.ENG.2017.03.020
Design of Photobioreactors for Mass Cultivation of Photosynthetic Organisms
a Qingdao Institute of Bioenergy and Bioprocess Technology, Chinese Academy of Sciences, Qingdao 266101, China
b Key Laboratory of Green Process and Engineering, Institute of Process Engineering, Chinese Academy of Sciences, Beijing 100190, China
c Nanomaterials Center, School of Chemical Engineering and Australian Institute for Bioengineering and Nanotechnology, The University of Queensland, Queensland 4072, Australia
Next Previous
Abstract
Photosynthetic microorganisms are important bioresources for producing desirable and environmentally benign products, and photobioreactors (PBRs) play important roles in these processes. Designing PBRs for photocatalysis is still challenging at present, and most reactors are designed and scaled up using semi-empirical approaches. No appropriate types of PBRs are available for mass cultivation due to the reactors’ high capital and operating costs and short lifespan, which are mainly due to a current lack of deep understanding of the coupling of light, hydrodynamics, mass transfer, and cell growth in efficient reactor design. This review provides a critical overview of the key parameters that influence the performance of the PBRs, including light, mixing, mass transfer, temperature, pH, and capital and operating costs. The lifespan and the costs of cleaning and temperature control are also emphasized for commercial exploitation. Four types of PBRs—tubular, plastic bag, column airlift, and flat-panel airlift reactors are recommended for large-scale operations. In addition, this paper elaborates the modeling of PBRs using the tools of computational fluid dynamics for rational design. It also analyzes the difficulties in the numerical simulation, and presents the prospect for mechanism-based models.
Keywords
Photobioreactor ; Solar energy ; Photosynthesis ; Hydrodynamics ; Flashing-light effect
Figures

Fig. 1
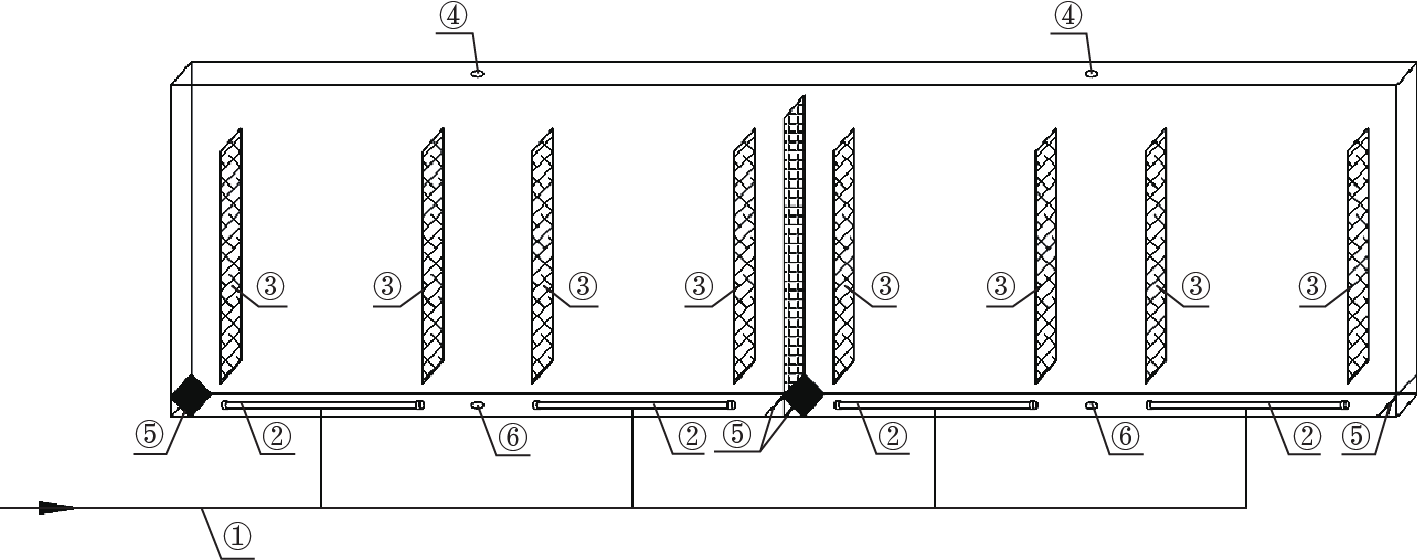
Fig. 2
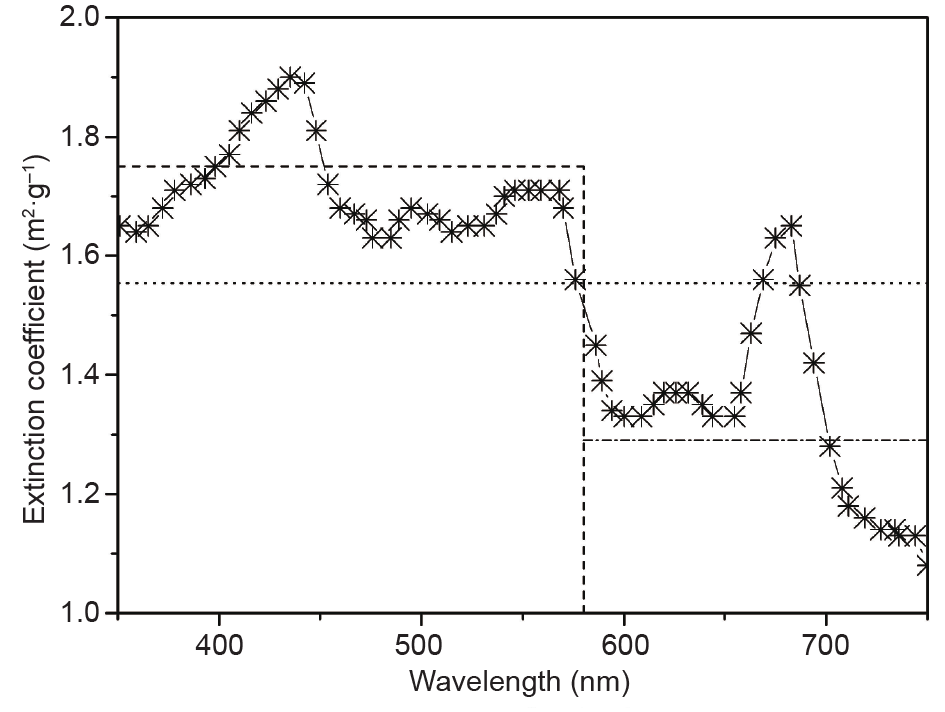
Fig. 3
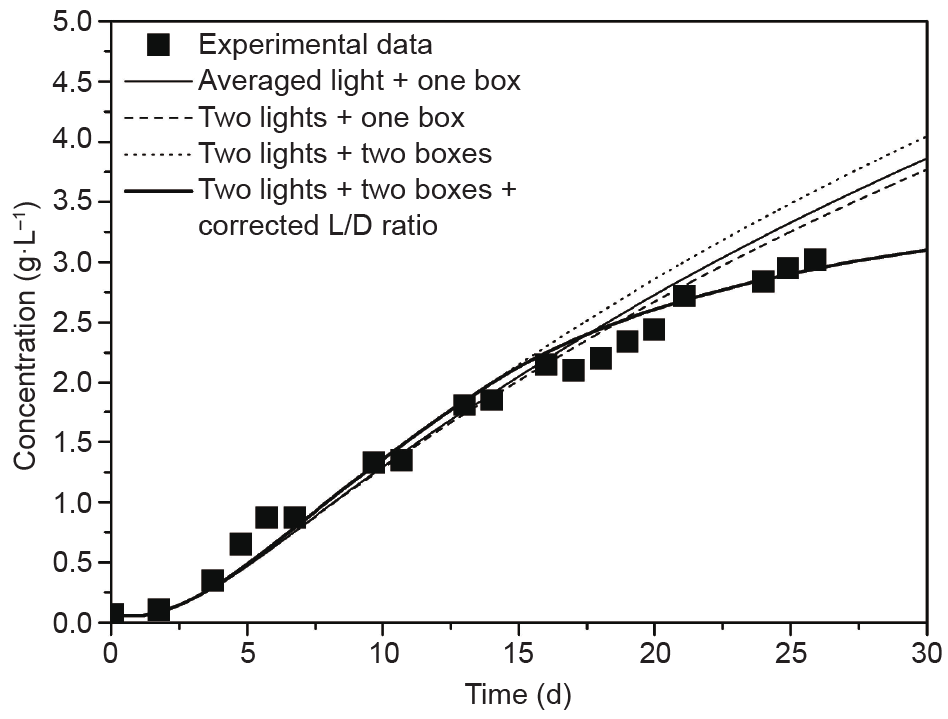
Fig. 4
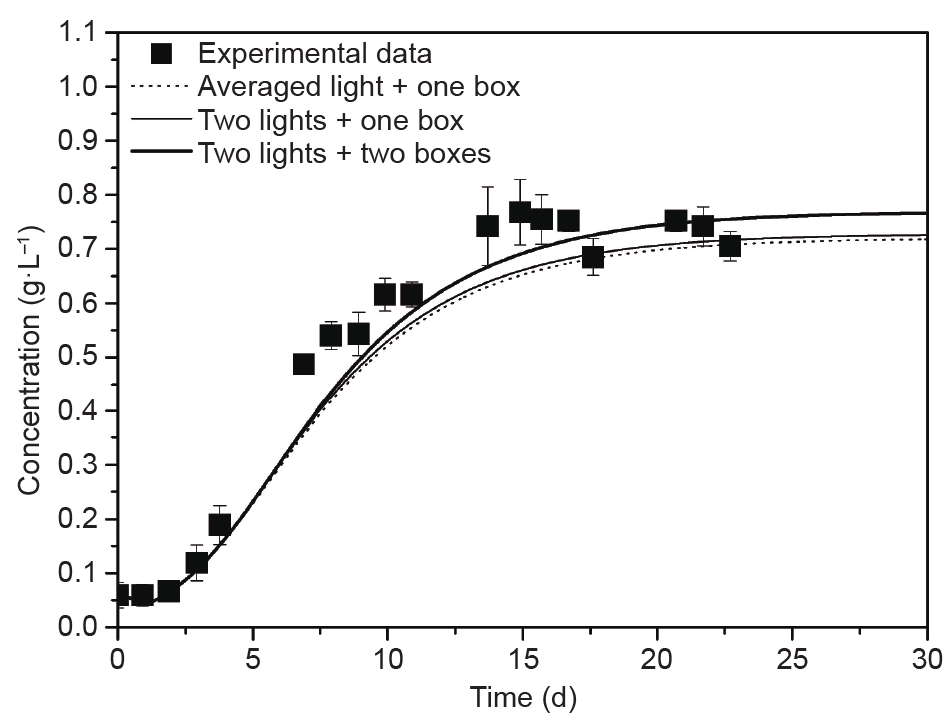
Fig. 5
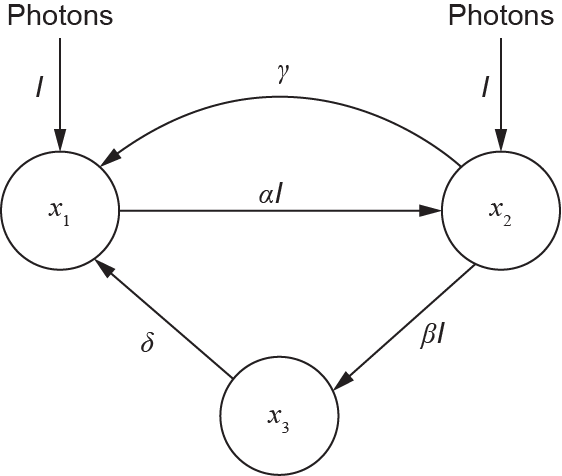
Fig. 6
References
[ 1 ] Knuckey RM, Brown MR, Robert R, Frampton DMF. Production of microalgal concentrates by flocculation and their assessment as aquaculture feeds. Aquacult Eng 2006;35(3):300–13 link1
[ 2 ] Takache H, Pruvost J, Marec H. Investigation of light/dark cycles effects on the photosynthetic growth of Chlamydomonas reinhardtiiin conditions representative of photobioreactor cultivation. Algal Res 2015;8:192–204 link1
[ 3 ] Shi XM, Chen F. High-yield production of lutein by the green microalga Chlorella protothecoidesin heterotrophic fed-batch culture. Biotechnol Prog 2002;18(4):723–7 link1
[ 4 ] Solimeno A, Samsó R, Uggetti E, Sialve B, Steyer JP, Gabarró A, et al.New mechanistic model to simulate microalgae growth. Algal Res 2015;12:350–8 link1
[ 5 ] Takache H, Christophe G, Cornet JF, Pruvost J. Experimental and theoretical assessment of maximum productivities for the microalgae Chlamydomonas reinhardtiiin two different geometries of photobioreactors. Biotechnol Prog 2010;26(2):431–40.
[ 6 ] Yan N, Fan C, Chen Y, Hu Z. The potential for microalgae as bioreactors to produce pharmaceuticals. Int J Mol Sci 2016;17(6):962 link1
[ 7 ] Del Campo JA, García-González M, Guerrero MG. Outdoor cultivation of microalgae for carotenoid production: Current state and perspectives. Appl Microbiol Biotechnol 2007;74(6):1163–74 link1
[ 8 ] Wang B, Lan CQ. Microalgae for biofuel production and CO2 sequestration. Hauppauge: Nova Science Publishers; 2010.
[ 9 ] Wang B, Lan CQ. Biomass production and nitrogen and phosphorus removal by the green alga Neochloris oleoabundans in simulated wastewater and secondary municipal wastewater effluent. Bioresour Technol 2011;102(10):5639–44 link1
[10] Abu-Ghosh S, Fixler D, Dubinsky Z, Iluz D. Continuous background light significantly increases flashing-light enhancement of photosynthesis and growth of microalgae. Bioresour Technol 2015;187:144–8 link1
[11] Batan LY, Graff GD, Bradley TH. Techno-economic and Monte Carlo probabilistic analysis of microalgae biofuel production system. Bioresour Technol 2016;219:45–52 link1
[12] Rodionova MV, Poudyal RS, Tiwari I, Voloshin RA, Zharmukhamedov SK, Nam HG, et al.Biofuel production: Challenges and opportunities. Int J Hydrogen Energy 2016;42(12):8450–61 link1
[13] Wang B, Lan CQ, Horsman M. Closed photobioreactors for production of microalgal biomasses. Biotechnol Adv 2012;30(4):904–12 link1
[14] Voloshin RA, Rodionova MV, Zharmukhamedov SK, Veziroglu TN, Allakhverdiev SI. Review: Biofuel production from plant and algal biomass. Int J Hydrogen Energy 2016;41(39):17257–73 link1
[15] Raeesossadati MJ, Ahmadzadeh H, McHenry MP, Moheimani NR. CO2 bioremediation by microalgae in photobioreactors: Impacts of biomass and CO2 concentrations, light, and temperature. Algal Res 2014;6(Part A):78–85.
[16] Posten C. Design principles of photo-bioreactors for cultivation of microalgae. Eng Life Sci 2009;9(3):165–77 link1
[17] Richmond A. Principles for attaining maximal microalgal productivity in photobioreactors: An overview. Hydrobiologia 2004;512(1):33–7 link1
[18] Huang Q, Liu T, Yang J, Yao L, Gao L. Evaluation of radiative transfer using the finite volume method in cylindrical photoreactors. Chem Eng Sci 2011;66(17):3930–40 link1
[19] Oncel S, Vardar Sukan F. Comparison of two different pneumatically mixed column photobioreactors for the cultivation of Artrospira platensis (Spirulina platensis). Bioresour Technol 2008;99(11):4755–60 link1
[20] Degen J, Uebele A, Retze A, Schmid-Staiger U, Trösch W. A novel airlift photobioreactor with baffles for improved light utilization through the flashing light effect. J Biotechnol 2001;92(2):89–94 link1
[21] Ogbonna J, Ichige E, Tanaka H. Interactions between photoautotrophic and heterotrophic metabolism in photoheterotrophic cultures of Euglena gracilis. Appl Microbiol Biotechnol 2002;58(4):532–8 link1
[22] Zittelli GC, Rodolfi L, Tredici MR. Mass cultivation of Nannochloropsissp. in annular reactors. J Appl Phycol 2003;15(2):107–14 link1
[23] Ranjbar R, Inoue R, Katsuda T, Yamaji H, Katoh S. High efficiency production of astaxanthin in an airlift photobioreactor. J Biosci Bioeng 2008;106(2):204–7 link1
[24] Rubio FC, Fernández FGA, Pérez JAS, Camacho FG, Grima EM. Prediction of dissolved oxygen and carbon dioxide concentration profiles in tubular photobioreactors for microalgal culture. Biotechnol Bioeng 1999;62(1):71–86 link1
[25] Ugwu C, Ogbonna J, Tanaka H. Improvement of mass transfer characteristics and productivities of inclined tubular photobioreactors by installation of internal static mixers. Appl Microbiol Biotechnol 2002;58(5):600–7 link1
[26] Harker M, Tsavalos AJ, Young AJ. Autotrophic growth and carotenoid production of Haematococcus pluvialis in a 30 liter air-lift photobioreactor. J Ferment Bioeng 1996;82(2):113–8 link1
[27] Kaewpintong K, Shotipruk A, Powtongsook S, Pavasant P. Photoautotrophic high-density cultivation of vegetative cells of Haematococcus pluvialis in airlift bioreactor. Bioresour Technol 2007;98(2):288–95 link1
[28] Hu Q, Guterman H, Richmond A. A flat inclined modular photobioreactor for outdoor mass cultivation of photoautotrophs. Biotechnol Bioeng 1996;51(1):51–60 link1
[29] Zhang T. Dynamics of fluid and light intensity in mechanically stirred photobioreactor. J Biotechnol 2013;168(1):107–16 link1
[30] Ogbonna JC, Soejima T, Tanaka H. An integrated solar and artificial light system for internal illumination of photobioreactors. J Biotechnol 1999;70(1–3):289–97 link1
[31] Richmond A, Boussiba S, Vonshak A, Kopel R. A new tubular reactor for mass production of microalgae outdoors. J Appl Phycol 1993;5(3):327–32 link1
[32] Hall DO, Acién Fernández FG, Guerrero EC, Rao KK, Grima EM. Outdoor helical tubular photobioreactors for microalgal production: Modeling of fluid-dynamics and mass transfer and assessment of biomass productivity. Biotechnol Bioeng 2003;82(1):62–73 link1
[33] Contreras A, García F, Molina E, Merchuk JC. Interaction between CO2-mass transfer, light availability, and hydrodynamic stress in the growth of Phaeodactylum tricornutum in a concentric tube airlift photobioreactor. Biotechnol Bioeng 1998;60(3):317–25 link1
[34] Watanabe Y, Saiki H. Development of a photobioreactor incorporating Chlorella sp. for removal of CO2 in stack gas. Energy Convers Manage 1997;38(Suppl):S499–503 link1
[35] Pruvost J, Pottier L, Legrand J. Numerical investigation of hydrodynamic and mixing conditions in a torus photobioreactor. Chem Eng Sci 2006;61(14):4476–89 link1
[36] Chetsumon A, Umeda F, Maeda I, Yagi K, Mizoguchi T, Miura Y. Broad spectrum and mode of action of an antibiotic produced by Scytonema sp. TISTR 8208 in a seaweed-type bioreactor. Appl Biochem Biotechnol 1998;70(1):249–56 link1
[37] Kumar K, Dasgupta CN, Nayak B, Lindblad P, Das D. Development of suitable photobioreactors for CO2 sequestration addressing global warming using green algae and cyanobacteria. Bioresour Technol 2011;102(8):4945–53 link1
[38] Moheimani NR, Isdepsky A, Lisec J, Raes E, Borowitzka MA. Coccolithophorid algae culture in closed photobioreactors. Biotechnol Bioeng 2011;108(9):2078–87 link1
[39] Singh RN, Sharma S. Development of suitable photobioreactor for algae production—A review. Renew Sustain Energy Rev 2012;16(4):2347–53 link1
[40] Sforza E, Simionato D, Giacometti GM, Bertucco A, Morosinotto T. Adjusted light and dark cycles can optimize photosynthetic efficiency in algae growing in photobioreactors. PLoS One 2012;7(6):e38975 link1
[41] Guo X, Yao L, Huang Q. Aeration and mass transfer optimization in a rectangular airlift loop photobioreactor for the production of microalgae. Bioresour Technol 2015;190:189–95 link1
[42] Melis A. Solar energy conversion efficiencies in photosynthesis: Minimizing the chlorophyll antennae to maximize efficiency. Plant Sci 2009;177(4):272–80 link1
[43] Yang Q, Pehkonen SO, Ray MB. Evaluation of three different lamp emission models using novel application of potassium ferrioxalate actinometry. Ind Eng Chem Res 2004;43(4):948–55 link1
[44] Merchuk JC, Wu X. Modeling of photobioreactors: Application to bubble column simulation. J Appl Phycol 2003;15(2):163–9 link1
[45] Bitog JP, Lee IB, Lee CG, Kim KS, Hwang HS, Hong SW, et al.Application of computational fluid dynamics for modeling and designing photobioreactors for microalgae production: A review. Comput Electron Agric 2011;76(2):131–47 link1
[46] Iluz D, Abu-Ghosh S. A novel photobioreactor creating fluctuating light from solar energy for a higher light-to-biomass conversion efficiency. Energy Convers Manage 2016;126:767–73 link1
[47] Zhang K, Kurano N, Miyachi S. Outdoor culture of a cyanobacterium with a vertical flat-plate photobioreactor: Effects on productivity of the reactor orientation, distance setting between the plates, and culture temperature. Appl Microbiol Biotechnol 1999;52(6):781–6 link1
[48] Zhang K, Miyachi S, Kurano N. Photosynthetic performance of a cyanobacterium in a vertical flat-plate photobioreactor for outdoor microalgal production and fixation of CO2. Biotechnol Lett 2001;23(1):21–6 link1
[49] Zemke PE, Sommerfeld MR, Hu Q. Assessment of key biological and engineering design parameters for production of Chlorella zofingiensis (Chlorophyceae) in outdoor photobioreactors. Appl Microbiol Biotechnol 2013;97(12):5645–55 link1
[50] Shang H, Scott JA, Shepherd SH, Ross GM. A dynamic thermal model for heating microalgae incubator ponds using off-gas. Chem Eng Sci 2010;65(16):4591–7 link1
[51] Anjos M, Fernandes BD, Vicente AA, Teixeira JA, Dragone G. Optimization of CO2 bio-mitigation by Chlorella vulgaris. Bioresour Technol 2013;139:149–54 link1
[52] Carvalho AP, Meireles LA, Malcata FX. Microalgal reactors: A review of enclosed system designs and performances. Biotechnol Prog 2006;22(6):1490–506 link1
[53] Acién Fernández FG, Fernández Sevilla JM, Molina Grima E. Photobioreactors for the production of microalgae. Rev Environ Sci Biotechnol 2013;12(2):131–51 link1
[54] Huang Q, Zhang W, Zhang G. Airlift loop reactors. In: Yang C, Mao ZS. Numerical simulation of multiphase reactors with continuous liquid phase. London: Elsevier Academic Press; 2014. p. 153–230.
[55] Xue S, Zhang Q, Wu X, Yan C, Cong W. A novel photobioreactor structure using optical fibers as inner light source to fulfill flashing light effects of microalgae. Bioresour Technol 2013;138:141–7 link1
[56] Grobbelaar JU. The influence of light/dark cycles in mixed algal cultures on their productivity. Bioresour Technol 1991;38(2–3):189–94 link1
[57] García-Camacho F, Sánchez-Mirón A, Molina-Grima E, Camacho-Rubio F, Merchuck JC. A mechanistic model of photosynthesis in microalgae including photoacclimation dynamics. J Theor Biol 2012;304:1–15 link1
[58] Vejrazka C, Janssen M, Streefland M, Wijffels RH. Photosynthetic efficiency of Chlamydomonas reinhardtii in flashing light. Biotechnol Bioeng 2011;108(12):2905–13 link1
[59] Abu-Ghosh S, Fixler D, Dubinsky Z, Iluz D. Flashing light in microalgae biotechnology. Bioresour Technol 2016;203:357–63 link1
[60] Vejrazka C, Streefland M, Wijffels RH, Janssen M. The role of an electron pool in algal photosynthesis during sub-second light-dark cycling. Algal Res 2015;12:43–51 link1
[61] Vejrazka C, Janssen M, Streefland M, Wijffels RH. Photosynthetic efficiency of Chlamydomonas reinhardtii in attenuated, flashing light. Biotechnol Bioeng 2012;109(10):2567–74 link1
[62] Kliphuis AMJ, de Winter L, Vejrazka C, Martens DE, Janssen M, Wijffels RH. Photosynthetic efficiency of Chlorella sorokiniana in a turbulently mixed short light-path photobioreactor. Biotechnol Prog 2010;26(3):687–96 link1
[63] Zijffers JWF, Schippers KJ, Zheng K, Janssen M, Tramper J, Wijffels RH. Maximum photosynthetic yield of green microalgae in photobioreactors. Mar Biotechnol 2010;12(6):708–18 link1
[64] Huang J, Li Y, Wan M, Yan Y, Feng F, Qu X, et al.Novel flat-plate photobioreactors for microalgae cultivation with special mixers to promote mixing along the light gradient. Bioresour Technol 2014;159:8–16 link1
[65] Zhu J, Rong J, Zong B. Factors in mass cultivation of microalgae for biodiesel. Chin J Catal 2013;34(1):80–100 link1
[66] Katsuda T, Shiraishi H, Ishizu N, Ranjbar R, Katoh S. Effect of light intensity and frequency of flashing light from blue light emitting diodes on astaxanthin production by Haematococcus pluvialis. J Biosci Bioeng 2008;105(3):216–20 link1
[67] Wu X, Merchuk JC. A model integrating fluid dynamics in photosynthesis and photoinhibition processes. Chem Eng Sci 2001;56(11):3527–38 link1
[68] Grobbelaar JU. Factors governing algal growth in photobioreactors: The “open” versus “closed” debate. J Appl Phycol 2009;21(5):489–92 link1
[69] Chisti Y. Biodiesel from microalgae. Biotechnol Adv 2007;25(3):294–306 link1
[70] Ugwu CU, Aoyagi H, Uchiyama H. Influence of irradiance, dissolved oxygen concentration, and temperature on the growth of Chlorella sorokiniana. Photosynthetica 2007;45(2):309–11 link1
[71] Pegallapati AK, Nirmalakhandan N. Internally illuminated photobioreactor for algal cultivation under carbon dioxide-supplementation: Performance evaluation. Renew Energy 2013;56:129–35 link1
[72] Chiu SY, Kao CY, Chen CH, Kuan TC, Ong SC, Lin CS. Reduction of CO2 by a high-density culture of Chlorella sp. in a semicontinuous photobioreactor. Bioresour Technol 2008;99(9):3389–96 link1
[73] Xing Z, Zong X, Pan J, Wang L. On the engineering part of solar hydrogen production from water splitting: Photoreactor design. Chem Eng Sci 2013;104:125–46 link1
[74] Singh SP, Singh P. Effect of temperature and light on the growth of algae species: A review. Renew Sustain Energy Rev 2015;50:431–44 link1
[75] Ugwu CU, Aoyagi H. Influence of shading inclined tubular photobioreactor surfaces on biomass productivity of C. sorokiniana.Photosynthetica 2008;46(2):283–5 link1
[76] Prakash J, Pushparaj B, Carlozzi P, Torzillo G, Montaini E, Materassi R. Microalgal biomass drying by a simple solar device. Int J Solar Energy 1997;18(4):303–11 link1
[77] Carlozzi P, Pushparaj B, Degl’Innocenti A, Capperucci A. Growth characteristics of Rhodopseudomonas palustriscultured outdoors, in an underwater tubular photobioreactor, and investigation on photosynthetic efficiency. Appl Microbiol Biotechnol 2006;73(4):789–95 link1
[78] Watanabe Y, de la Noüe J, Hall DO. Photosynthetic performance of a helical tubular photobioreactor incorporating the cyanobacterium Spirulina platensis. Biotechnol Bioeng 2011;47(2):261–9 link1
[79] Georgianna DR, Mayfield SP. Exploiting diversity and synthetic biology for the production of algal biofuels. Nature 2012;488(7411):329–35 link1
[80] Acién Fernández FG, García Camacho F, Sánchez Pérez JA, Fernández Sevilla JM, Molina Grima E. Modeling of biomass productivity in tubular photobioreactors for microalgal cultures: Effects of dilution rate, tube diameter, and solar irradiance. Biotechnol Bioeng 1998;58(6):605–16 link1
[81] Berberoglu H, Pilon L, Melis A. Radiation characteristics of Chlamydomonas reinhardtii CC125 and its truncated chlorophyll antenna transformants tla1, tlaX and tla1-CW+. Int J Hydrogen Energy 2008;33(22):6467–83 link1
[82] Vasumathi KK, Premalatha M, Subramanian P. Parameters influencing the design of photobioreactor for the growth of microalgae. Renew Sustain Energy Rev 2012;16(7):5443–50 link1
[83] Fan LH, Zhang YT, Zhang L, Chen HL. Evaluation of a membrane-sparged helical tubular photobioreactor for carbon dioxide biofixation by Chlorella vulgaris. J Membr Sci 2008;325(1):336–45 link1
[84] Li S. The dynamics of CO2 fixation by microalgae [dissertation]. Qingdao: Qingdao Institute of Bioenergy and Bioprocess Technology, Chinese Academy of Sciences; 2013.
[85] Iglesias-RodrÍGuez MD, Nimer NA, Merrett MJ. Carbon dioxide-concentrating mechanism and the development of extracellular carbonic anhydrase in the marine picoeukaryote Micromonas pusilla. New Phytol 1998;140(4):685–90 link1
[86] Moheimani NR, Borowitzka MA. Increased CO2 and the effect of pH on growth and calcification of Pleurochrysis carterae and Emiliania huxleyi (Haptophyta) in semicontinuous cultures. Appl Microbiol Biotechnol 2011;90(4):1399–407 link1
[87] Luo HP, Kemoun A, Al-Dahhan MH, Sevilla JMF, Sánchez JLG, Camacho FG, et al.Analysis of photobioreactors for culturing high-value microalgae and cyanobacteria via an advanced diagnostic technique: CARPT. Chem Eng Sci 2003;58(12):2519–27 link1
[88] Perner-Nochta I, Posten C. Simulations of light intensity variation in photobioreactors. J Biotechnol 2007;131(3):276–85 link1
[89] Berenguel M, Rodrı?guez F, Acién FG, Garcı?a JL. Model predictive control of pH in tubular photobioreactors. J Process Contr 2004;14(4):377–87 link1
[90] Sierra E, Acién FG, Fernández JM, García JL, González C, Molina E. Characterization of a flat plate photobioreactor for the production of microalgae. Chem Eng J 2008;138(1–3):136–47 link1
[91] Ugwu C, Ogbonna J, Tanaka H. Design of static mixers for inclined tubular photobioreactors. J Appl Phycol 2003;15(2):217–23 link1
[92] Vree JH, Bosma R, Janssen M, Barbosa MJ, Wijffels RH. Comparison of four outdoor pilot-scale photobioreactors. Biotechnol Biofuels 2015;8(1):215 link1
[93] Chen CY, Chang JS, Chang HY, Chen TY, Wu JH, Lee WL. Enhancing microalgal oil/lipid production from Chlorella sorokiniana CY1 using deep-sea water supplemented cultivation medium. Biochem Eng J 2013;77:74–81 link1
[94] Abomohra AEF, El-Sheekh M, Hanelt D. Pilot cultivation of the chlorophyte microalga Scenedesmus obliquus as a promising feedstock for biofuel. Biomass Bioenergy 2014;64:237–44 link1
[95] Kim ZH, Park H, Hong SJ, Lim SM, Lee CG. Development of a floating photobioreactor with internal partitions for efficient utilization of ocean wave into improved mass transfer and algal culture mixing. Bioprocess Biosyst Eng 2016;39(5):713–23 link1
[96] Huang J, Ying J, Fan F, Yang Q, Wang J, Li Y. Development of a novel multi-column airlift photobioreactor with easy scalability by means of computational fluid dynamics simulations and experiments. Bioresour Technol 2016;222:399–407 link1
[97] Soman A, Shastri Y. Optimization of novel photobioreactor design using computational fluid dynamics. Appl Energy 2015;140:246–55 link1
[98] Huang Q, Yao L, Guo X. A device consisted of airlift flat-panel photobioreactors for the cultivation of photosynthetic microorganism. China patent, Appl Num: 201510478717.1. 2015.
[99] Dasgupta CN, Jose Gilbert J, Lindblad P, Heidorn T, Borgvang SA, Skjanes K, et al.Recent trends on the development of photobiological processes and photobioreactors for the improvement of hydrogen production. Int J Hydrogen Energy 2010;35(19):10218–38 link1
[100] Meiser A, Schmid-Staiger U, Trösch W. Optimization of eicosapentaenoic acid production by Phaeodactylum tricornutumin the flat panel airlift (FPA) reactor. J Appl Phycol 2004;16(3):215–25 link1
[101] Huang J, Feng F, Wan M, Ying J, Li Y, Qu X, et al.Improving performance of flat-plate photobioreactors by installation of novel internal mixers optimized with computational fluid dynamics. Bioresour Technol 2015;182:151–9 link1
[102] Wheaton ZC, Krishnamoorthy G. Modeling radiative transfer in photobioreactors for algal growth. Comput Electron Agric 2012;87:64–73 link1
[103] Pareek VK, Cox SJ, Brungs MP, Young B, Adesina AA. Computational fluid dynamic (CFD) simulation of a pilot-scale annular bubble column photocatalytic reactor. Chem Eng Sci 2003;58(3–6):859–65 link1
[104] Denny F, Scott J, Pareek V, Ding Peng G, Amal R. CFD modelling for a TiO2-coated glass-bead photoreactor irradiated by optical fibres: Photocatalytic degradation of oxalic acid. Chem Eng Sci 2009;64(8):1695–706 link1
[105] Wu X, Merchuk JC. Simulation of algae growth in a bench-scale bubble column reactor. Biotechnol Bioeng 2002;80(2):156–68 link1
[106] Wu X, Merchuk JC. Simulation of algae growth in a bench scale internal loop airlift reactor. Chem Eng Sci 2004;59(14):2899–912 link1
[107] Janssen M, Janssen M, de Winter M, Tramper J, Mur LR, Snel J, et al.Efficiency of light utilization of Chlamydomonas reinhardtii under medium-duration light/dark cycles. J Biotechnol 2000;78(2):123–37 link1
[108] Suh IS, Lee SB. A light distribution model for an internally radiating photobioreactor. Biotechnol Bioeng 2003;82(2):180–9 link1
[109] Benson BC, Gutierrez-Wing MT, Rusch KA. The development of a mechanistic model to investigate the impacts of the light dynamics on algal productivity in a hydraulically integrated serial turbidostat algal reactor (HISTAR). Aquacult Eng 2007;36(2):198–211 link1
[110] Bosma R, van Zessen E, Reith JH, Tramper J, Wijffels RH. Prediction of volumetric productivity of an outdoor photobioreactor. Biotechnol Bioeng 2007;97(5):1108–20 link1
[111] Elyasi S, Taghipour F. Simulation of UV photoreactor for degradation of chemical contaminants: Model development and evaluation. Environ Sci Technol 2010;44(6):2056–63 link1
[112] Li D, Xiong K, Li W, Yang Z, Liu C, Feng X, et al.Comparative study in liquid-phase heterogeneous photocatalysis: Model for photoreactor scale-up. Ind Eng Chem Res 2010;49(18):8397–405 link1
[113] Pottier L, Pruvost J, Deremetz J, Cornet JF, Legrand J, Dussap CG. A fully predictive model for one-dimensional light attenuation by Chlamydomonas reinhardtii in a torus photobioreactor. Biotechnol Bioeng 2005;91(5):569–82 link1
[114] Santos PD, Lani A. An object-oriented implementation of a parallel Monte Carlo code for radiation transport. Comput Phys Commun 2016;202:233–61 link1
[115] Rochatte V, Dahi G, Eskandari A, Dauchet J, Gros F, Roudet M, et al.Radiative transfer approach using Monte Carlo method for actinometry in complex geometry and its application to Reinecke salt photodissociation within innovative pilot-scale photo(bio)reactors. Chem Eng J 2017;308:940–53 link1
[116] Trujillo Francisco J, Lee Ivy AL, Hsu CH, Safinski T, Adesina Adesoji A. Hydrodynamically-enhanced light intensity distribution in an externally-irradiated novel aerated photoreactor: CFD simulation and experimental studies. Int J Chem React Eng 2008;6(1):A58.
[117] Ben Salah M, Askri F, Slimi K, Ben Nasrallah S. Numerical resolution of the radiative transfer equation in a cylindrical enclosure with the finite-volume method. Int J Heat Mass Transfer 2004;47(10–11):2501–9 link1
[118] Pareek V, Chong S, Tadé M, Adesina AA. Light intensity distribution in heterogenous photocatalytic reactors. Asia-Pac J Chem Eng 2008;3(2):171–201 link1
[119] Chai JC, Lee HS, Patankar SV. Finite volume method for radiation heat transfer. J Thermophys Heat Transfer 1994;8(3):419–25 link1
[120] van de Hulst HC. Light scattering by small particles. 2nd ed. New York: Dover Publications; 1981.
[121] Huang Q, Yao L, Liu T, Yang J. Simulation of the light evolution in an annular photobioreactor for the cultivation of Porphyridium cruentum. Chem Eng Sci 2012;84:718–26 link1
[122] Berberoglu H, Yin J, Pilon L. Light transfer in bubble sparged photobioreactors for H2 production and CO2 mitigation. Int J Hydrogen Energy 2007;32(13):2273–85 link1
[123] Modest MF. Radiative heat transfer. 2nd edition. New York: Academic Press; 2003.
[124] Pruvost J, Cornet JF, Legrand J. Hydrodynamics influence on light conversion in photobioreactors: An energetically consistent analysis. Chem Eng Sci 2008;63(14):3679–94 link1
[125] Huang Q, Yang C, Yu G, Mao ZS. 3-D simulations of an internal airlift loop reactor using a steady two-fluid model. Chem Eng Technol 2007;30(7):870–9 link1
[126] Huang Q, Yang C, Yu G, Mao ZS. CFD simulation of hydrodynamics and mass transfer in an internal airlift loop reactor using a steady two-fluid model. Chem Eng Sci 2010;65(20):5527–36 link1
[127] Huang Q, Zhang W, Yang C. Modeling transport phenomena and reactions in a pilot slurry airlift loop reactor for direct coal liquefaction. Chem Eng Sci 2015;135:441–51 link1
[128] Lee E, Jalalizadeh M, Zhang Q. Growth kinetic models for microalgae cultivation: A review. Algal Res 2015;12:497–512 link1
[129] Pruvost J, Legrand J, Legentilhomme P, Muller-Feuga A. Simulation of microalgae growth in limiting light conditions: Flow effect. AIChE J 2002;48(5):1109–20 link1
[130] Pruvost J, Legrand J, Legentilhomme P, Muller-Feuga A. Lagrangian trajectory model for turbulent swirling flow in an annular cell: Comparison with residence time distribution measurements. Chem Eng Sci 2002;57(7):1205–15 link1
[131] Rosello Sastre R, Csögör Z, Perner-Nochta I, Fleck-Schneider P, Posten C. Scale-down of microalgae cultivations in tubular photo-bioreactors—A conceptual approach. J Biotechnol 2007;132(2):127–33 link1
[132] Muller-Feuga A, Le Guédes R, Pruvost J. Benefits and limitations of modeling for optimization of Porphyridium cruentum cultures in an annular photobioreactor. J Biotechnol 2003;103(2):153–63 link1
[133] Eilers PHC, Peeters JCH. A model for the relationship between light intensity and the rate of photosynthesis in phytoplankton. Ecol Modell 1988;42(3–4):199–215 link1
[134] Podola B, Li T, Melkonian M. Porous substrate bioreactors: A paradigm shift in microalgal biotechnology? Trends Biotechnol 2017;35(2):121–32 link1











 京公网安备 11010502051620号
京公网安备 11010502051620号




