《1. Introduction》
1. Introduction
Thermal burn trauma causes very complex tissue injury, altering biomolecular elements, structures, and secondary degradation processes. Cell viability depends upon proper structure and function of each of these constituents. When cells are subjected to temperature histories that exceed the threshold of the intramolecular bonds, cell plasma membranes, and proteins denature, ultimately resulting in injury [1]. Unfortunately, burn injury is a common occurrence. It is estimated that over six million people around the world suffer a significant burn each year [2]. It is generally believed that thermal injuries are irreversible. However, eukaryotic cells have a certain capacity for self-repair that is both constituent and inducible, permitting living systems to adapt to harsh environments by the upregulation of molecular repair mechanisms. By investigating and understanding the biochemical mechanisms of cell repair, it is possible to develop strategies to mimic these processes. Here we discuss how molecular engineering science can be used to more effectively preserve functional tissue recovery following thermal burn injury.
《2. Molecular basis of thermal injury》
2. Molecular basis of thermal injury
A thermal burn results from the exposure of tissues, most commonly the skin, to supra-physiological temperatures for durations sufficient to alter molecular structures and tissue viability. In clinical practice today, the goal of resuscitation and wound care therapies for burn victims is to preserve organ function and limit the spread of damage. While this approach saves many lives, it does not rescue thermally damaged components of tissue. The result is a degree of permanent functional loss dependent upon the tissues involved and the extent of the injury [2].
Typically, burn injuries presenting in clinics are spatially non-uniform, reflecting the gradient in temperature history and variance in the molecular composition and tissue densities [3]. The fractional extent of thermal injury at any point in the tissue directly coincides with the temperature history and the conformational stability of the cellular and extracellular biomacromolecules or the supramolecular assembly of biomacromolecules. The conformational stabilities vary over a wide range from lipid bilayer membranes to nucleic acids.
The cell plasma membrane is composed of amphiphilic proteins and phospholipids held together by entropic forces of hydration. As the temperature increases above the 41−44 °C range, the kinetic energy of the molecules in the cell plasma membrane exceeds the hydration energy barrier that holds phospholipids in the membrane as a supramolecular assembly [4,5]. In effect, the heated membrane forms defects and becomes indiscriminately permeable, or “porous” to ions. Once permeabilized, the cell attempts to maintain the transmembrane ionic gradients by increasing the activity of cellular energy pumps, resulting in the rapid depletion of cellular energy charge. Unless the membrane defects are sealed before the degradation processes exceed certain limits, the permeabilized cell will lose viability. Because the cell plasma membrane is one of the most thermally labile cell structures, yet is critical for cell viability, tissue loss from thermal injury is heavily mediated by cell membrane disruption [4].
Protein structure tends to be more stable against thermal fluctuation than the lipid assembly in the cell plasma membrane due to stronger bonding forces. The melting temperatures of common structural proteins actin (intracellular) and collagen (extracellular), measured by near-equilibrium healing kinetics, have been reported at 67 °C and 58 °C respectively, well above the melting temperature of the cell plasma membrane. However, the melting temperature of protein denaturation is highly dependent on tissue density, macromolecular crowding, and the temperature history. Once thermal denaturation occurs, the exposed hydrophobic regions interact, resulting in biomolecular aggregation and alteration of both cell water structure and metabolic dynamics. Unless corrected, cell death followed by tissue necrosis will inexorably occur.
《3. Natural cellular injury repair processes》
3. Natural cellular injury repair processes
The capability to survive and adapt to environmental stress is a fundamental characteristic of successful living systems. By exposure to sub-lethal thermal injury, eukaryotic cells can be “preconditioned” to tolerate higher levels of thermal injury within hours after the initial exposure [6]. This preconditioning involves restoration of both the cell plasma membrane and protein structures. Cells are intrinsically capable of “sealing” plasma membrane defects by transporting cytoplasmic vesicles up ionic gradients, toward the membrane defect, where it fuses with the membrane [7]. The normal recycling of membrane constituents leads to the replacement of the membrane with new membrane over time. These processes involved in sealing defects can be upregulated by repeated damage, which is the typical paradigm for achieving injury tolerance.
Similarly, there exist mechanisms for the repair or removal of denatured proteins. In response to supra-physiological thermal history, the accumulation of denatured cellular proteins triggers upregulation of the genes responsible for natural molecular chaperone biosynthesis. These chaperones, initially called heat shock proteins (HSPs), exhibit several functions that enhance cell survival after injury, including the “chaperoning” or guided refolding of thermally damaged proteins. One molecular mechanism of HSP action pertains to binding to exposed hydrophobic regions of denatured proteins and entropically driving disaggregation and refolding. Disaggregation is the first step that allows proteins to spontaneously refold into their native, more energetically favorable, conformation [6]. Some chaperones requiring adenosine triphosphate (ATP) will actively assist protein refolding through a range of physicochemical mechanisms. Harnessing these intrinsic repair mechanisms could greatly increase viability.
《4. Applications of molecular engineering》
4. Applications of molecular engineering
Many physicochemical processes involved in cellular repair or adaptation can serve as therapeutic targets for molecular engineering. In fact, simple amphiphilic multiblock copolymers, such as poloxamers, have been effective in accelerating cell repair processes across various injury models. They have been used to reseal thermally injured cells and chaperone the functional recovery of heat denatured proteins (Figure 1) [8−12]. Even more dramatic examples include reports that human foreskin fibroblasts treated with poloxamers maintained their morphological integrity and contractile function after exposure to 45 °C for 1 h [8]. Corroborating in vivo studies have shown that the application of poloxamers to burn wounds increased blood flow [9,10] and reduced burn depth [9,11,12].
《Fig. 1》
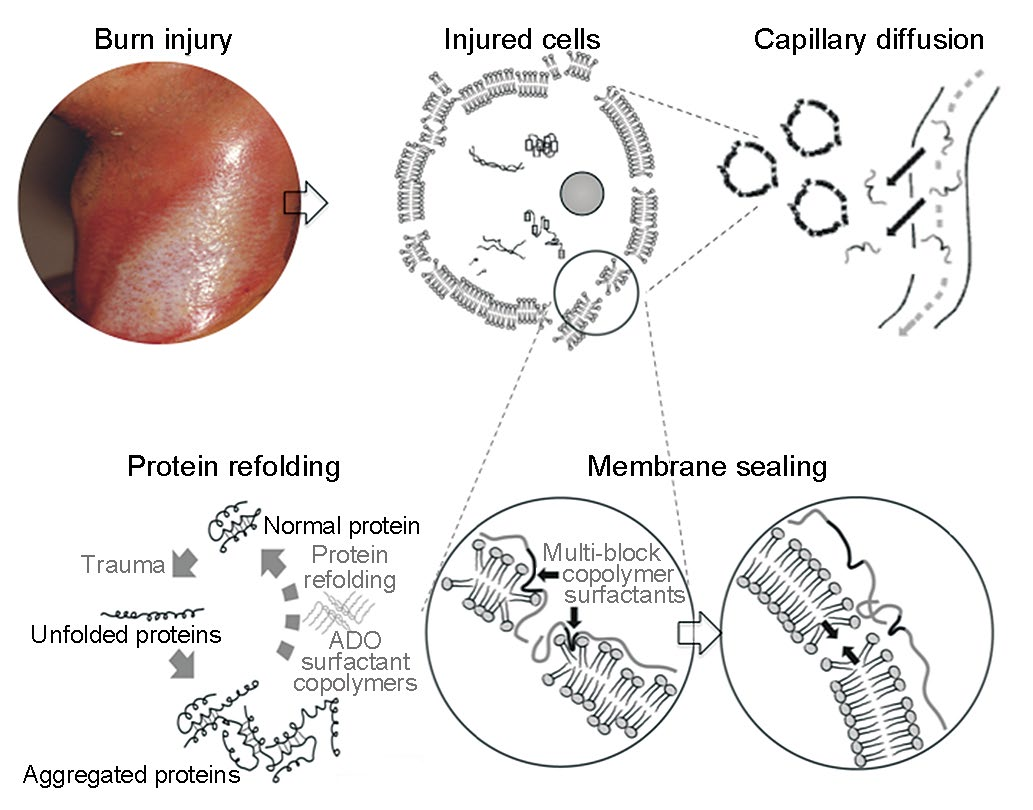
Fig.1 Illustration of the pharmaceutical action of small block copolymer surfactants in the treatment of a thermal skin burn wound.
Amphiphic block copolymers have been shown to faciliate the refolding and recovery of catalytic activity of thermally injured proteins [6,13]. However, copolymer design must match the denatured protein structure. Evidence to date suggests that the copolymer surfactant interacts with the exposed hydrophobic regions of denatured proteins, inhibiting or blocking intermolecular binding and subseqent aggregation. The net result is the facilitation of spontaneous refolding [13].
The primary objective of current clinical practice in burn care focuses on inhibiting secondary injury processes and replacing nonviable tissue. The next advance will most likely emerge from the molecular engineering of macromolecules that mimic natural chaperone function or catalzye enhanced chaperone functionality. Success in this therapeutic goal would result in tremendous benefit to the millions who suffer burn injuries every year.
《Compliance with ethics guidelines》
Compliance with ethics guidelines
Kristen Jakubowski, Michael Poellmann, and Raphael C. Lee declare that they have no conflict of interest or financial conflicts to disclose.











 京公网安备 11010502051620号
京公网安备 11010502051620号




