
《1. Introduction》
1. Introduction
Most big rivers on the earth have been intensively dammed for hydropower production, flood management, water supply, as well as navigation (see Fig. S1(a) in Appendix A), and the number of dams is still rapidly growing (see Fig. S1(b)). River damming not only alters the natural hydrologic regime [1], sediment flux [1,2] and water quality [3,4], but also results in undesirable ecological effects on fish community [5,6]. Dam operations affect the spawning activity of native fish species and thus damage their populations and communities [7,8], which has been reported on rivers around the world, such as the Madeira River [9], the Colorado River [10], and the Lancang–Mekong River [8]. Fish spawning activity is likely triggered by a wide range of environmental factors, including discharge [11], water temperature [12], dissolved oxygen [13], turbidity [14], and primary production [15]. Flow velocity is known particularly critical to the spawning of fishes delivering drifting eggs [13,16,17].
Chinese carps, which include silver carp (Hypophthalmichthys molitrix), grass carp (Ctenopharyngodon idellus), black carp (Mylopharyngodon piceus), and bighead carp (Aristichthys nobilis), play important roles in both cultivation and capture fishery in China [18]. In the 20th century, 60% of the entire catch of freshwater fishes in the Yangtze River was composed of Chinese carps [19]. However, in the last decades, due to the remarkable changes in hydrological conditions caused by the construction of the Three Gorges Dam, the spawning activities of the Chinese carp species have been severely impacted (see Fig. S1(c)). In 2003, the number of fish eggs and larvae was 10% of that in 2002, when the Three Gorges Reservoir (TGR) started operation [20,21].
In other places, Chinese carps have become one of the most disastrous invasive species (see Fig. S2(a)). Due to their biological attributes such as high fecundities and movement rates, the populations expanded rapidly, and successfully invaded into many rivers and lakes [22]. The invasion and population explosion of Chinese carps in the United States (see Figs. S1(b) and (c)) has caused serious decline of native fish populations [23]. In Australia, Chinese carps sustain high biomasses (up to 3144 kg·hm-2 ) in many rivers [24]. In China, many native or endemic species in plateau lakes have been destructively impacted by the introduction of bighead and silver carp [25], leading to irreversible ecological changes. Despite a variety of measures have been taken to control Chinese carps, even a highly risk idea is proposed to release virus to prevent the expansion of Chinese carp population in Australia [26,27], little effective outcome has been achieved.
Managing the reproduction, including spawning, fertilizing, hatching, and embryo development, could be an effective measure to sustain the population or control the invasion of Chinese carps. However, studies so far focus only on the spawning, which is reported to take place in large and turbid rivers with a minimum water temperature requirement of 18 °C [28] and specific flow velocity requirements [13,16,17]. No solid scientific supports are explored to establish a quantitative relationship between flow velocity and Chinese carp reproduction, considering both spawning and fertilizing, hatching, as well as larvae surviving.
In this paper, we took H. molitrix as the target species and used a physical model to test two hypotheses: ① There exists quantitative relationships between flow velocity and spawning activity, as well as embryo development of Chinese carp; ② there exists an optimal flow velocity balancing both the requirements of spawning activity and embryo development. Meanwhile, multiple-year field experiments on Chinese carp spawning were conducted to crossevidence the control experiments. The study provides a solid and fundamental basis to improving or impeding fish reproduction by adaptive management of hydraulic engineering structures in rivers.
《2. Material and methods》
2. Material and methods
All the experiments were conducted in the Dangtu Experimental Centre of Nanjing Hydraulic Research Institute, China, under a permission in accordance with the guidelines of the Institutional Animal Care and Use Committee (IACUC).
《2.1. Collection of fish eggs in the field experiments》
2.1. Collection of fish eggs in the field experiments
Samples of drifting eggs were collected at a fixed cross-section, which is near Yidu City (Fig. 1), in the main stream of Yangtze River during the field flow manipulation experiment from 2012 to 2016. Larvae and drifting eggs of Chinese carps were frequently monitored at this cross-section during the reproduction season. The reach from Yichang to Yangjiazui, which is about 10 km upstream of the cross-section, is reported to be the spawning ground [29]. The reach is nearly 1 km wide, and has neither erodible riffles nor narrow sections. These special morphological features create suitable flow conditions that benefit to Chinese carp spawning.
《Fig. 1》

Fig. 1. The studied river reach and its riverbed elevation as well as the cross-sections (CS1–CS4) for spawned egg sampling in the middle Yangtze River.
During the five-year field experiments, sampling started at midMay in each year, when water temperature was higher than the threshold of 18 °C. The sampling continued for two consecutive weeks until no egg was found. Fish eggs were collected from 8:00 to 9:00 and from 14:00 to 15:00 every day during the sampling period. A net (0.5 m diameter, 500 μm mesh) was used to collect drifting eggs. The net was suspended on the water surface and was oriented to upstream against the flow. A propeller-type current meter was used to measure the flow velocity simultaneously. The corresponding discharge (m3 ·s-1 ) at the upstream boundary and water level (m) at the downstream boundary of the reach were obtained from Changjiang Water Resources Commission.
The collected eggs were immediately examined. The eggs with a diameter of 4–5 mm were considered as potential Chinese carp eggs. Developmental stages and post-fertilization time at collection were both identified. The identified eggs were incubated in the laboratory until their species were confirmed, so that egg density (eggs per 1000 m3 ) of each sampling was finally estimated.
To obtain the flow field of the entire reach, a two-dimensional unsteady hydrodynamic model was developed. The model was calibrated and validated by the obtained data of discharge and water level as well as the measured cross-section velocity at four cross-sections of CS1–CS4 (Fig. 1). The details of model description, calibration and verification of the hydrodynamic model were presented in Appendix A Section 1. The calibrated model was then used to calculate the flow velocity of the entire river reach.
Based on the calculated flow velocity and the embryonic development time (drift time), the precise spawning sites in upstream were located by computing the drift distance away from the egg collection cross-section (see Appendix A Section 2). The corresponding flow velocities of each spawning event at the located sites were then extracted from the modeled flow field and were used for statistical analysis.
《2.2. Laboratory experiments》
2.2. Laboratory experiments
The lab experiment was conducted from May 15th to June 20th in 2017 and 2018, respectively. The experimental device was a large annular flume (Fig. 2(a)). Two submerged water impellers and four submerged pumps were installed to accelerate the flow of water. The middle of the straight sections was widened and deepened to facilitate the placement of the submerged water impeller, and fishing nets were installed on both sides to prevent the silver carps from entering into the impeller area. Twenty-two measuring points were arranged around the flume, and the velocity at 0.5 m depth was measured by propeller-typed current meter (LS20B, Nanjing Water Conservancy and Hydrology Automation Institute, China). There were six underwater cameras around the flume to observe the spawning behavior of H. molitrix. During the experiment, the water depth was set at 0.9 m and the water quality was consistent with that of the acclimation pond. Water quality parameters were measured using a multifunctional water quality analyzer (YSI 650MDS, YSI Inc., USA). The experiment designed six scenarios with different theoretically maximum velocities (see Table S1), and each scenario was performed twice in each year.
《Fig. 2》

Fig. 2. (a) The large annular flume with a depth of 1.8 m for spawning test, and (b) a small annular flume with a depth of 0.2 m for embryo experiment.
Sexually mature males (length: (37 ± 1.3) cm; weight: (3.6 ± 0.4) kg; number: 72) and females (length: (43 ± 2.1) cm, weight: (4.5 ± 0.6) kg; number: 72) of H. molitrix were obtained from the Chinese carps Hatchery Center in Huanggang City, Hubei Province, China. Before the experiments, they were placed in a net cage in an acclimation pond. A commercial fish diet was supplied to satiation twice a day. After seven days, six male and six female H. molitrix were randomly selected as a group for experiment. Before being moved into the annular flume, the selected H. molitrix were injected with luteinizing hormone-releasing hormone antagonist-2 (LHRHA2) once. The injection dosage was 2 μg·kg-1 body weight for female, and it was reduced half for male. The injection position was at the base of pectoral fin. They were tested for three days or until eggs were laid in each scenario. After each test, female H. molitrix were anesthetized using a sublethal dose of NaOH-neutralized ethyl 3-aminobenzoate methanesulfonate (MS-222) and dissected to confirm the number of spawning females according to ovarian saturation.
To test the effects of flow velocity on embryo development, a small annular flume was set up (Fig. 2(b)), which used small booster pumps to control the flow velocity. Three scenarios with designed maximum flow velocity of 0.4, 0.8, and 1.2 m·s-1 were tested. Each velocity treatment was repeated in triplicate. The total hatch ratio and the survival ratio of newly hatched larvae 24 h post-hatch were used to assess the embryo development. Water quality parameters were measured in the same way as above. In the small annular flumes, a constant water level was maintained and half of the water was replaced every day. Two hundred eggs were selected from the large flume to assess the development of embryos in each test if spawning happed. These tests were only conducted in June 2018.
To obtain the flow velocity of the entire annular flume, computational fluid dynamics (CFD) method was used in the study, which was implemented in CFX 11.0 (ANSYS, Inc., USA). The model was calibrated and validated by the observed data from the 22 measuring points. The details of the model description, calibration and verification were presented in Appendix A Section 3.
《2.3. Statistical analysis of experimental data》
2.3. Statistical analysis of experimental data
The fish egg data from lab and field experiments as well as the flow velocity data from hydrodynamic/CFX models were analyzed by one-way analysis of variance (ANOVA) to determine the effects of flow velocity on the spawning and embryo development of H. molitrix. Significance was analyzed using Duncan’s multiple range test and Tukey’s student range test at a significance (P) of 0.05. Data were expressed as mean ± standard error (SE). All analyses were conducted using SigmaPlot 11.0 (Systat Software, Inc., USA).
《3. Results》
3. Results
《3.1. Reproduction of H. molitrix in field experiments in the midYangtze River》
3.1. Reproduction of H. molitrix in field experiments in the midYangtze River
From the five-year field experiments, 20 spawning events (dashed boxes) were detected, among which 11 peak spawning events (higher dashed boxes) were identified (Fig. 3). For each spawning event, the duration of days (the width of each pink block in Fig. 3) was calculated. Then the averaged duration is obtained, which was about 4.5 days.
《Fig. 3》
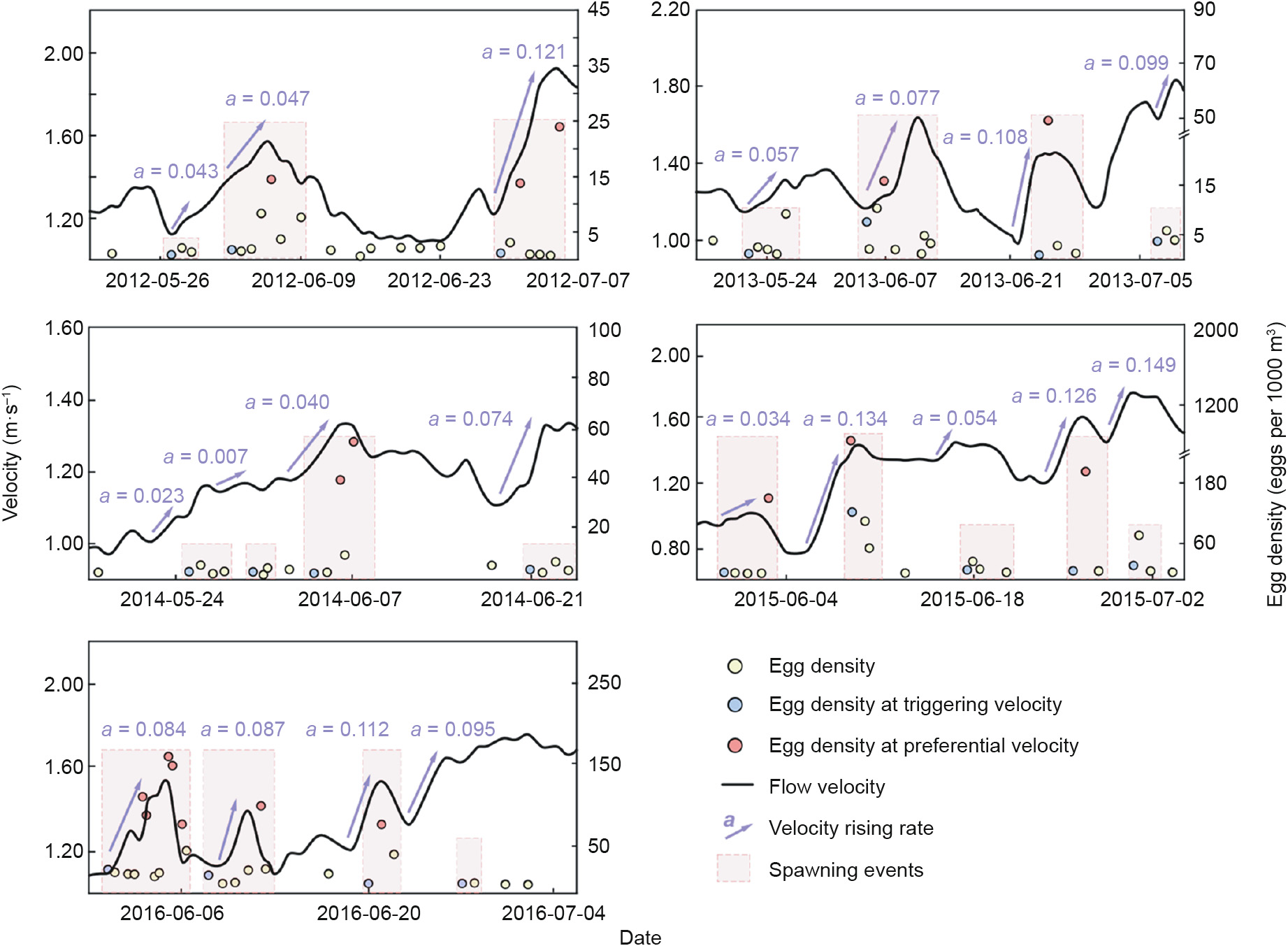
Fig. 3. Fish spawning events and the corresponding flow velocity as well as velocity rising rate during 2012 to 2016 in mid-Yangtze River in the field experiments. The black curves show the flow velocity at the spawning site. The dots are the observed egg density. The red dash lines indicate spawning events. The blue dots represent the egg density under triggering velocity, which is defined as the flow velocity when first spawning action took place in a spawning event. The red dots are the egg density under suitable velocity in which a prominent spawning occurred. The blue arrows indicate the velocity rising rate (a) during spawning events.
During the sampling periods, water temperatures were at suitable levels, ranging from 18.5 to 24.0 °C (see Table S2). Egg identification showed that about 55% of collected eggs belonged to silver carp, and 38% belonged to grass carp, and 5% belonged to black carp, and only 1% belonged to bighead carp. The results also indicated that the egg proportions of different Chinese carp species were similar every year, which meant there was no significant change in the proportion between the five years. The developmental stage of fish embryos was clarified to determine the drift time of eggs. The identified eggs were predominantly at last blastula stage, which accounted for 72.42% of the total, while 27.58% of the examined eggs were identified as the other developmental stages (see Table S3).
The calibration and validation results of hydrodynamic model were shown in Fig. S3. It was seen that the relative root mean square error for flow velocity and water level during validation ranged in 0.7%–4.1% and 0.15%–0.23%, respectively, implying that the modeled flow fields of the river reach were reliable. The determined spawning sites showed that the locations of spawning events were between Yanzhiba and Honghuatao (Fig. 4(a)), which were consistent with previous studies [30]. The corresponding flow velocities of each spawning event at the located sites were extracted from the hydrodynamic model results and were analyzed with the egg density data in Fig. 3. The statistical results (Fig. 4) showed that in the surveyed river reach, the flow velocity triggering H. molitrix to spawn was (1.30 ± 0.20) m·s-1 (Fig. 4(b)), and the preferential velocity for H. molitrix to spawn was (1.40 ± 0.23) m·s-1 (Fig. 4(c)).
《Fig. 4》

Fig. 4. (a) Flow velocity of the studied river reach and the located spawning sites of the spawning events; (b) triggering velocity (n = 20) and (c) preferential velocity (n = 11) in the mid-Yangtze River in the field experiments. SD: standard deviation; n: number of spawning events.
《3.2. Reproduction of H. molitrix in flume simulation experiments》
3.2. Reproduction of H. molitrix in flume simulation experiments
The flow fields of the large annular flume for different experiment scenarios were shown in Fig. 5. Since no spawning activity was observed for the experimental scenarios with a maximum velocity of 0.8 and 1.8 m·s-1 , the results were not taken for further analysis. The videos from the six underwater cameras (Fig. 2) showed that all the spawning activities (see Supplementary videos Breeding fish and Spawned eggs) happened in bend sections. Accordingly, the flow velocities at these bends were extracted from the CFD simulated flow fields for further analyses. The water quality conditions in the flume during the experiments were shown in Table S4.
《Fig. 5》

Fig. 5. Simulated flow field of the large annular flume under different experiment scenarios.
The ratio of spawned females to total tested females was plot with the extracted flow velocity for each experimental scenario (Fig. 6(a)). The ratio increased significantly with the increase in flow velocity until 1.4 m·s-1 , and then decreased sharply at 1.6 m·s-1 . The highest ratio of spawned female, which reached 67%, was observed at the flow velocity of 1.4 m·s-1 (Fig. 6(a)). However, the maximum fertilization ratio of eggs occurred at the flow velocity of 1.2 m·s-1 (Fig. 6(b)), and it was significantly higher than those at the velocity of 1.0 and 1.6 m·s-1 . There was no significant difference in fertilization ratio between 1.2 and 1.4 m·s-1 .
The hatch ratio (hatched eggs/total experimental eggs) of H. molitrix embryo declined with the increase in flow velocity (Fig. 6(c)). At the velocity of 1.2 m·s-1 , the hatch ratio was as low as 15%. The survival ratio of larvae in 24 h (survived larvae/total experimental eggs) showed a similar trend to the hatch ratio. The survival ratio was about 12% in static water and decreased to 4% at 1.2 m·s-1 . The low hatch ratio and survival ratio were mainly due to the low fertilization ratio of the experimental eggs, which were collected from the large flume. The high flow velocities in the large flume during spawning experiments caused low fertilization rate.
《Fig. 6》

Fig. 6. Reproduction of H. molitrix at different flow velocities in lab experiments. (a) The ratios of spawning females with flow velocity (n = 4); (b) fertilization ratios with flow velocity (n = 4); and (c) the hatch ratio of fertilized eggs and survival ratio of larvae (n = 3). The curves were fitted using one-dimensional Gaussian function.
《3.3. Consistency between lab and field experimental results》
3.3. Consistency between lab and field experimental results
In the flume experiments, the highest egg density appeared at the flow velocity of 1.4 m·s-1 and the lowest egg density was at the flow velocity of 1.6 m·s-1 (Fig. 7(a)). The analyses on the spawning event data from the field showed that a velocity range from 1.05 to 1.60 m·s-1 was suitable for H. molitrix to spawn in natural water bodies (Fig. 7(b)).
The egg density data from both lab and field experiments were normalized according to Vismara et al. [31]. The normalized spawning data (S) and the flow velocity (v) were fitted using Gaussian regression, which gave S = exp[(–v –1.3062)2 /0.0707] (R2 = 0.9289, P < 0.0001) for the flume experiments and S = exp[(–v – 1.3143)2 /0.0968] (R2 = 0.9828, P < 0.0001) for the field experiments (Fig. 7). The results from both lab and field experiments were highly consistent, which could be jointly written as S = exp[(–v – 1.31)2 /0.097]. Therefore, an optimal flow velocity of 1.31 m·s-1 was identified for the spawning of H. molitrix in rivers.
《Fig. 7》

Fig. 7. Fish reproduction against flow velocity. (a) Normalized egg density at different velocity level and its Gaussian fitting curve in the flume experiments. (b) The frequency histogram and the Gaussian fitting curve of spawning activities in natural spawning sites.
《3.4. Testing application of the study》
3.4. Testing application of the study
The practical testing of the results to the adaptive operation of the TGR for Chinese carps conservation showed promising prospects. Based on the requirements of minimum velocity limit and daily velocity rise and maximum velocity limit as well as the cross-section geometry of the Yidu spawning ground, the discharge was suggested to start from 12 500 m3 ·s-1 (with a flow velocity of 1.05 m·s-1 ) at initial day when a spawning event begins, and gradually increase to 18 600 m3 ·s-1 (with a flow velocity of 1.40 m·s-1 ) at day 4 when the spawning reaches the peak, and then quickly drop down to support hatching and larvae survival (Fig. 8(a) and Appendix A Section 4). This discharge process is repeated during Chinese carps spawning season, which is from middle May to middle June in the downstream of TGR. During the testing from 2017 to 2019, eggs were measured following the same procedures as the egg collections during the field experiments to evaluate the efficiency. The results showed that the suggested operation in TGR has continuously made significant achievements in increasing the spawned eggs of Chinese carps (Fig. 8(b)).
《Fig. 8》

Fig. 8. (a) The suggested flow process for engineering application to the TGR operation for improving Chinese carps reproduction in the downstream of Three Gorges Dam. (b) The changes of spawned eggs of Chinese carps in the downstream of TGR since the implementation of the flow process. Data is from China Three Gorges Corporation and China National Environmental Monitoring Center.
《4. Discussion》
4. Discussion
We quantified the effects of flow velocity on the spawning and embryo development of H. molitrix, a species delivering drifting eggs, through both field and lab experiments. Our research offered direct evidence that a certain flow velocity was a prerequisite for silver carp to spawn. In the flume experiments, the percentage of successfully spawned female silver carp increased with the increase in flow velocity, and reached the maximum at a flow velocity of 1.4 m·s-1 (Fig. 3). However, when flow velocity was as high as 1.6 m·s-1 , the spawning activity was severely inhibited, which was due to the excessive physical exertion for H. molitrix to keep swimming. In the natural spawning sites of Yangtze River, silver fish eggs were observed at a wider velocity range from 1.05 to 1.60 m·s-1 . The wider range of preferential flow velocity in natural water bodies could be attributed to the complicated hydrological conditions and habitat characteristics. Besides preferential flow velocity for H. molitrix spawning, triggering velocity was another important factor. No spawning activity was observed in flume at a velocity of 0.8 m·s-1 , and the determined triggering velocity was about 1.0 m·s-1 (Fig. 4). In the natural river reach, spawning was observed at a velocity range from 0.90 to 1.90 m·s-1 (Fig. 4). The required flow velocity for spawning differs largely among different fish species. Female Atlantic salmon constructs spawning redds in areas with an averaged flow velocity of 0.53 m·s-1 [32], while the spawning of medaka (Oryzias latipes) decreases under flowing water and recovers when water flow stops [33]. Our research demonstrated that though silver carp is widely found living and growing in relatively lentic habitats [34], a certain velocity of water flow is required for its reproduction.
Flow velocity was not only a trigger for the spawning activity of H. molitrix, but also influenced the fertilization ratio of eggs. The highest fertilization ratio was observed at 1.2 m·s-1 , but there was no significant difference in fertilization ratios between 1.2 and 1.4 m·s-1 (Fig. 3). Due to the external fertilization character of silver carps, the sperm-egg fusion happens in water through mixing effects induced by water flow. Because the swimming ability of sperm was limited both temporally and spatially [35], a flow velocity higher than 1.6 m·s-1 caused a rapid decrease in fertilization due to the failure of encounters between sperm and eggs. After fertilization, eggs need to be hatched under a certain flow velocity, as the eggs of silver carp are semi-buoyant and must remain suspended during the development of early life stage. Garcia et al. [36] has studied the sedimentation of silver carp eggs at different flow velocities and found that 0.2 m·s-1 could support the suspension of eggs. However, in this study, both the hatch ratio and survival ratio of newly hatched larvae of silver carp decreased significantly with the increase in flow velocity. It showed that lentic water was more favorable to the early life stage development. According to Murphy and Jackson [37], flow velocity as low as 0.15 to 0.25 m·s-1 could be sufficient to suspend the silver carp eggs in four tributaries of the Great Lakes. The early life stages of fish, especially during the rapidly growing embryo and larval stages, are most suitable for assessing environmental pressure, as the larvae and juvenile fish are more prone to environmental stress than adults [38]. Small changes in environmental factors may cause considerable variation in phenotype, resulting in a significant impact on survival and development in later ontogeny [39]. Excessive flow velocity causes mechanical damage to eggs [36], which could be the main reason for the inhibition of both egg hatching and larval survival. After fertilization, eggs floated downstream, and embryonic development happened in relatively slow currents. Otherwise, even though the higher flow velocity could stimulate spawning of fish, they did not lead to increased recruitment because larval survival was low under rapid flows [40]. Besides the major function of water flow to maintain the eggs in suspension, water exchange and dissolved oxygen content is also increased at higher flow velocity, which can benefit to hatching. In our flume experiment, the water depth was not as large as in natural water bodies and the egg densities were low as well, therefore the oxygen conditions were not the limiting factor for the development of early life stage. In natural water bodies, the preferential flow velocity for egg hatching of H. molitrix needs to be further determined.
It has been well known that the breeding performance of teleosts is regulated by pituitary hormones, especially gonadotropin (GtH), which are regulated by gonadotropin releasing hormone (GnRH) [41]. Environmental conditions, such as a rising discharge, long photoperiod and warm temperatures, are thought to be important factors controlling the reproduction cycles in teleosts. The activity of GnRH is observed to be up-regulated under the environmental factors that stimulate gonadal activity [42]. A rising discharge has been reported to be the primary cue for spawning of many teleosts, including silver carp. Unlike common carps, for example Cyprinus carpio, silver carp is unable to spawn naturally in lentic water, because of the stagnation of ovary development in mature females of silver carps. The artificial injection of LHRHA2 has been demonstrated to be effective to induce spawning of silver carp [43]. In our experiment, LHRH-A2 was applied to investigate the necessity of flow velocity in the induction of spawning of silver carp. A flow velocity of 0.8 m·s-1 could not trigger the spawning activity of silver carp even after the injection of LHRHA2, and spawning occurred only when the flow velocity reached 1.0 m·s-1 .
In China, the Yangtze River is the main habitat for silver carp. There are several dams in the upper reach of the river, which is known to have caused the loss of spawning grounds for Chinese carps [29]. The establishment of new spawning grounds will highly affect the maintenance of the population. This study offered the explicit range of flow velocity required for spawning of silver carp, which provided an essential clue to dam operation for Chinese carp conservation. In contrast to the population depression in Yangtze River, Chinese carps are causing serious ecological disasters in many countries as an invasive species. Silver carp has been introduced to at least 88 countries or territories for aquaculture or for controlling algal blooms or as invasive species, and more than a third of these introductions have already established selfsustaining populations [44]. The introductions have caused severe consequences due to the competitive advantage of silver carp over native species and profound influence on plankton communities in the long term [45]. It is reported that the biomass of silver carp comprised 51% of total fish collections in the La Grange Reach of the Illinois River in 2008 [46]. High phenotypic plasticity in breeding traits is the reason for the successful establishment of the new populations [47]. Meanwhile, a series of physical and non-physical barriers, including dam blocking [48], electric dispersal barriers [49], acoustic stimuli barriers [50], or biological methods, including employment of koi herpesvirus [51] and the introduction of female-specific-sterility and female-specific-lethality individuals [52], have been applied to suppress the spreading of silver carp populations. However, these measures have not successfully solved the problem, and even worse, some of them have induced unexpected ecological risks [26,27]. Since spawning and early life stage development play the primary role in the establishment of an invasive species, blocking the spawning process will be of critical importance to the control of invasion. The flow velocity requirement is rigid in the spawning of silver carp, but is much more flexible during the rest of the life stages [35]. Therefore, manipulating flow velocity by dams can be a cost-effective way to suppress the invasion of H. molitrix. Given that most of big rivers in the world are heavily dammed, the findings provided a fundamental basis to manipulate flow velocities during reproduction season to enhance (1.2 ≤ v ≤ 1.4 m·s-1 ) or control (v < 0.8 m·s-1 or v > 1.8 m·s-1 ) the population of carps by adapting operations of reservoirs in dammed rivers.
《5. Conclusions》
5. Conclusions
Recent studies have overwhelmingly stressed the negative consequences of dam construction, notably the effects on fish. Chinese carps play an important role in food security and protein accessibility in China; however, their natural population in Yangtze River has declined sharply due to the remarkable changes in hydrological conditions incurred by river damming. Flow velocity is known a primary factor to fish early life development, which is decisive to fish population. We for the first time quantitatively identified the preferential velocity range for Chinese carps reproduction, considering both spawning requirements and embryo developments, through lab and field experiments. Our findings provide the fundamental knowledge to manage Chinese carp natural populations through non-engineering approaches, such as adaptive flow regulation by dams in rivers. Future study will focus on multiobjective optimization of cascade reservoirs with consideration of hydrological uncertainties.
《Acknowledgements》
Acknowledgements
This work is supported by the National Key Research and Development Program of China (2016YFC0502205) and the National Natural Science Foundation of China (51425902).
《Authors’ contribution》
Authors’ contribution
Qiuwen Chen, Jianyun Zhang and Lei Chen designed the study, supervised the experiments, analyzed the data, and finished manuscript. Lei Tang, Jun Wang, Yunqing Lin, Yong Gao, Wei Jiang and Yuxin Zhang conducted the experiments. Kangle Mo and Yuchen Chen analyzed the data and prepared the figures. All authors contributed to the preparation of the manuscript.
《Compliance with ethics guidelines》
Compliance with ethics guidelines
Qiuwen Chen, Jianyun Zhang, Yuchen Chen, Kangle Mo, Jun Wang, Lei Tang, Yuqing Lin, Lei Chen, Yong Gao, and Wei Jiang declare that they have no conflict of interest or financial conflicts to disclose.
《Appendix A. Supplementary data》
Appendix A. Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.eng.2020.02.010.











 京公网安备 11010502051620号
京公网安备 11010502051620号




