

《1 前言》
1 前言
在一般情况下,硬骨鱼类排出体外的成熟卵子处于第二次减数分裂中期。鱼类的这种繁殖特征为鱼类的细胞工程育种,例如:多倍体诱导、雄核发育诱导以及雌核发育诱导。人工诱导雌核发育是一种快速固定母本性状、生产单性化苗种以及建立纯系的有效手段。根据原理的不同,人工诱导雌核发育又可进一步被分为减数分裂雌核发育(Meiogynogenesis)和有 丝分 裂雌 核发 育(Mitogynogenesis)。减数分裂雌核发育比有丝分裂雌核发育更易操作,有更高的诱导率,目前已经成功地应用于上百种鱼类[1~3] 。
牙鲆(Paralichthys olivaceus)是一种重要经济鱼类,雌性个体生长速度明显比雄性快。中国水产科学研究院北戴河中心实验站运用雌核发育诱导技术,选育的全雌牙鲆新品种“北鲆1号”和“北鲆2 号”,具有雌性比例高、生长快、规格整齐、容易饲养等特点。关于牙鲆雌核发育的研究,已经有众多论文报导[4~6] 。王桂兴等[7] 采用微卫星技术,研究了牙鲆连续两代减数分裂雌核发育家系的遗传特征,结果显示,第二代减数分裂雌核发育家系的纯合度、个体间平均相似度以及亲子间的平均相似度要略高于第一代雌核发育家系,显著高于对照家系。
本研究在王桂兴等[7] 技术的基础上,对性成熟的第二代雌核发育个体再度诱导减数分裂雌核发育,获得三个连续三代减数分裂雌核发育家系,并利用 30对微卫星引物分析了该三个家系的纯合度和遗传相似度,旨在探讨牙鲆连续多代雌核发育过程对等位基因纯合进程的影响,为牙鲆通过连续多代减数分裂雌核发育方法建立纯系以及优良品种选育提供相应的数据支持。
《2 材料和方法》
2 材料和方法
《2.1 连续三代减数分裂雌核发育家系的构建》
2.1 连续三代减数分裂雌核发育家系的构建
本实验在中国水产科学研究院北戴河中心实验站开展。2007年,以真鲷精子作为异源精子,经紫外线照射灭活,激活一尾雌性野生牙鲆所产卵子,并在激活后3 min,施以3 ℃,45 min的冷休克处理进行减数分裂雌核发育诱导,获得第一代减数分裂雌核发育家系(Meio-G1)。2009 年,利用 MeioG1内发育成熟的一尾个体再度诱导减数分裂雌核发育,获得连续第二代减数分裂雌核发育家系(Meio-G2)。2014 年,以 Meio-G2 家系内三尾性成熟个体进行减数分裂雌核发育诱导,获得了三个连续三代减数分裂雌核发育家系(Meio-G3-1、MeioG3-2、Meio-G3-3)。
《2.2 样品采集与基因组DNA提取》
2.2 样品采集与基因组DNA提取
采集作为母本的三尾 Meio-G2 的腹面胸鳍和 Meio- G3- 1、Meio- G3- 2、Meio- G3- 3 家系仔鱼各 30尾,置于100 %酒精中固定,-20 ℃保存。基因组 DNA提取采用传统的酚-氯仿抽提法进行[7] 。
《2.3 微卫星引物》
2.3 微卫星引物
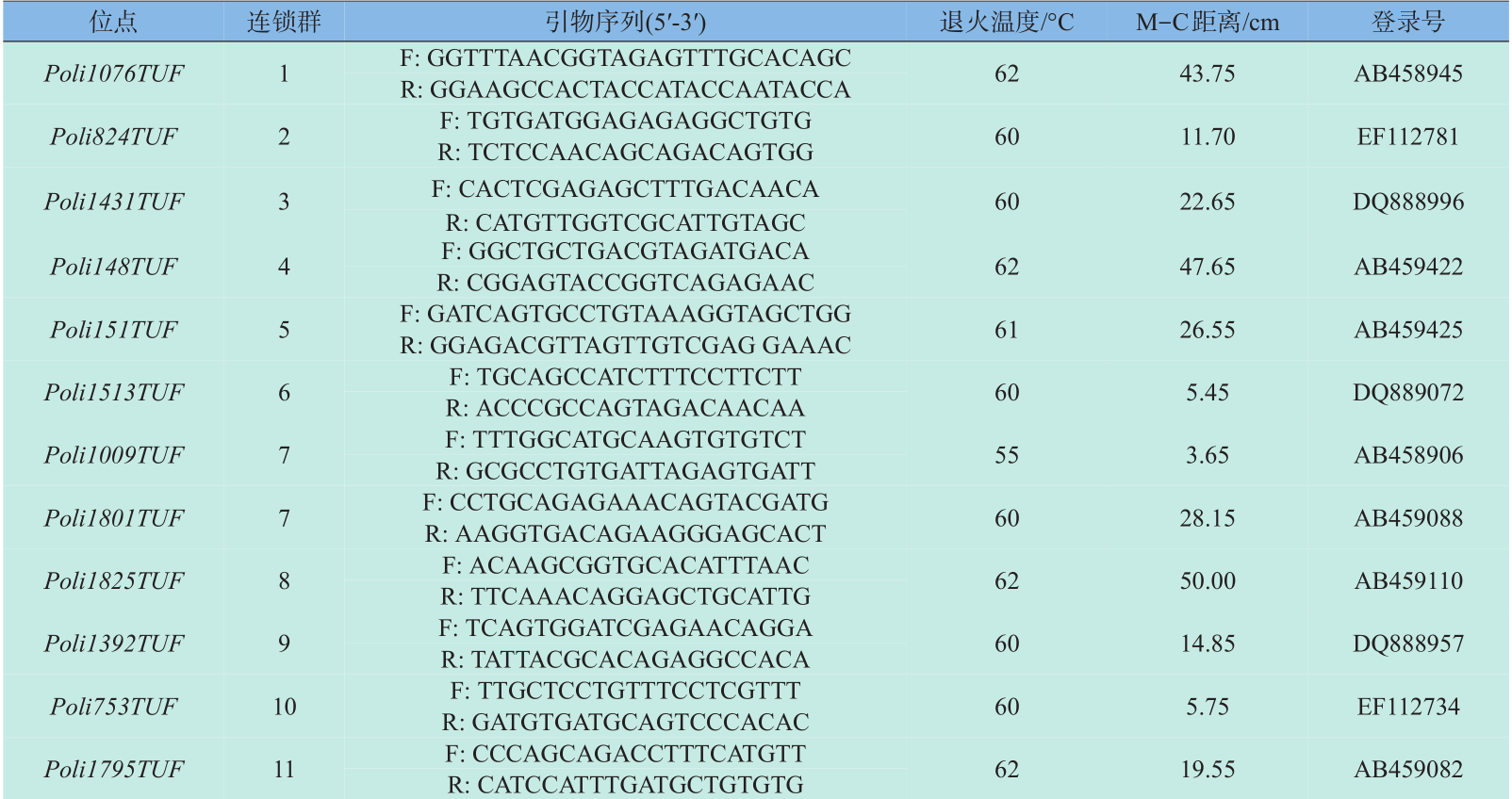
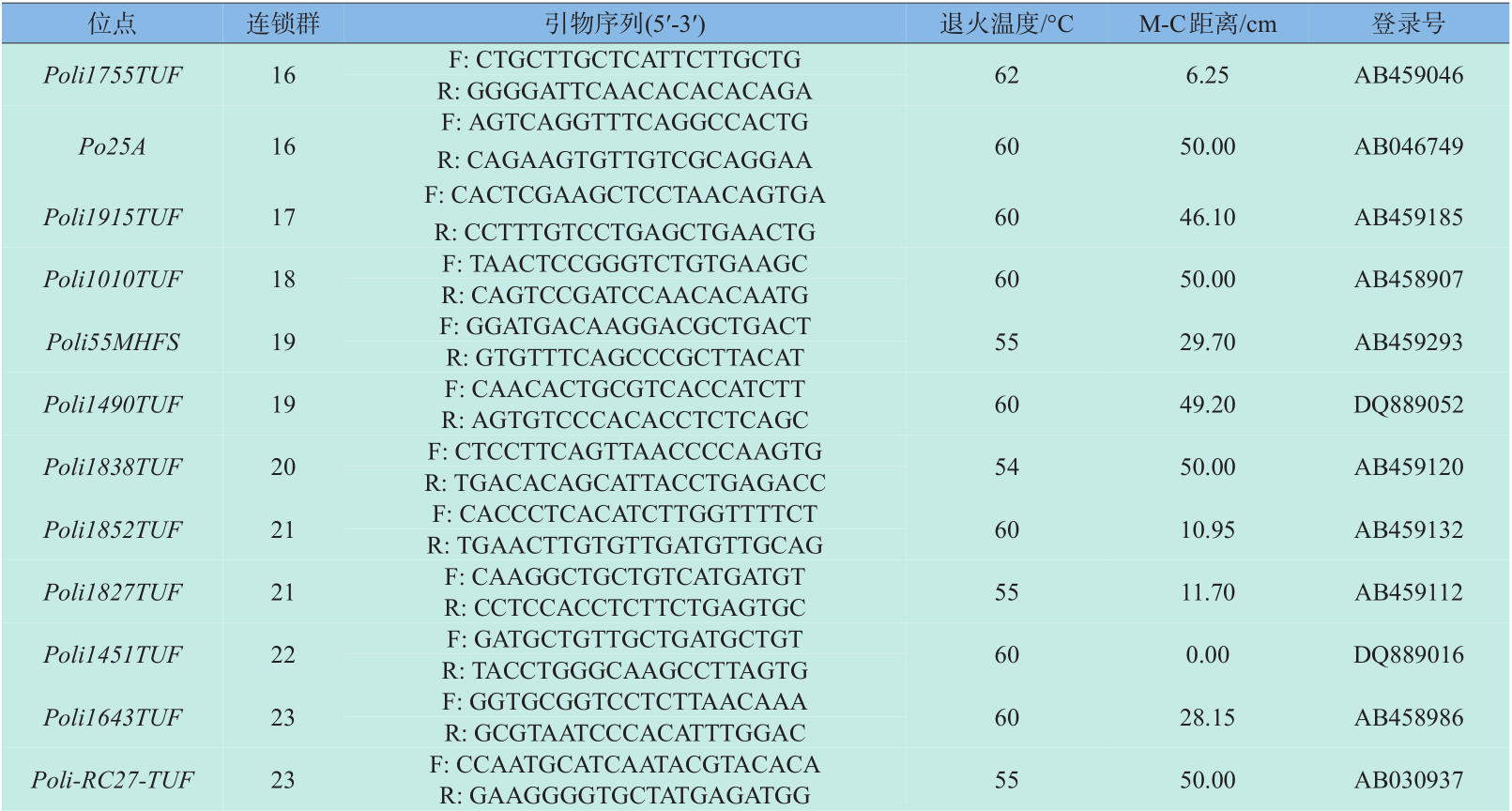
微卫星引物全部选自Coimbra等[8] 绘制的牙鲆第二代遗传连锁图谱,根据刘永新等[9] 的实验结果,选取重组率高、中、低的引物各10对,所选引物覆盖牙鲆24个连锁群。引物由上海生工生物工程技术服务有限公司合成。引物序列、退火温度、M-C 距离和Genbank登录号见表1。
《表1》
表1 30对微卫星标记位点引物序列、退火温度、M-C距离和Genbank登录号
Table 1 Primer sequences,annealing temperatures,M-C distance and accession number in GenBank of 30 microsatellite loci in the Paralichthys olivaceus

续表

《2.4 聚合酶链式反应(PCR)扩增及电泳》
2.4 聚合酶链式反应(PCR)扩增及电泳
PCR 反应体系采用 15 μL,包括:10×Buffer 2.5 μL、Mg2+ (25 mmol/L)1 μL、dNTPs(2 mmol/L) 1 μL、上下游引物(10 mmol/L)各0.6 μL、DNA模板 1 μL(40~50 ng)、Taq DNA 聚合 酶 1 U,加适 量 ddH2O。反应程序为 94 ℃预变性 4 min;共 25 个循环包括94 ℃变性30 s,退火30 s,72 ℃延伸30 s;最后 72 ℃延伸 8 min,PCR 扩增在 PE9700 型 PCR 仪上进行。PCR产物用8 %非变性聚丙烯酰胺凝胶电泳,银染显色后用HP Scanjet G4010扫描仪成像,用 Gel-Pro Analyzer4.5凝胶分析软件对电泳图像进行数据采集和分析。
《2.5 统计指标》
2.5 统计指标
等位基因数(number of alleles,A)、观测杂合度(Ho)、纯合度(homozygosity)、遗传相似系数(genetic similarity index,GSI)等遗传参数均由 popgene (ver.3.2)计算获得。重组率和近交系数由以下公式计算

近交系数
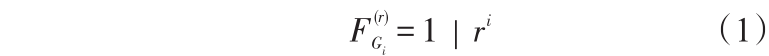
式(1)中,F 为第i 世代的近交系数;Gi 为第i 代雌核发育家系;r 为微卫星位点的重组率。
《3 结果与分析》
3 结果与分析
《3.1 PCR扩增及基因纯合度》
3.1 PCR扩增及基因纯合度
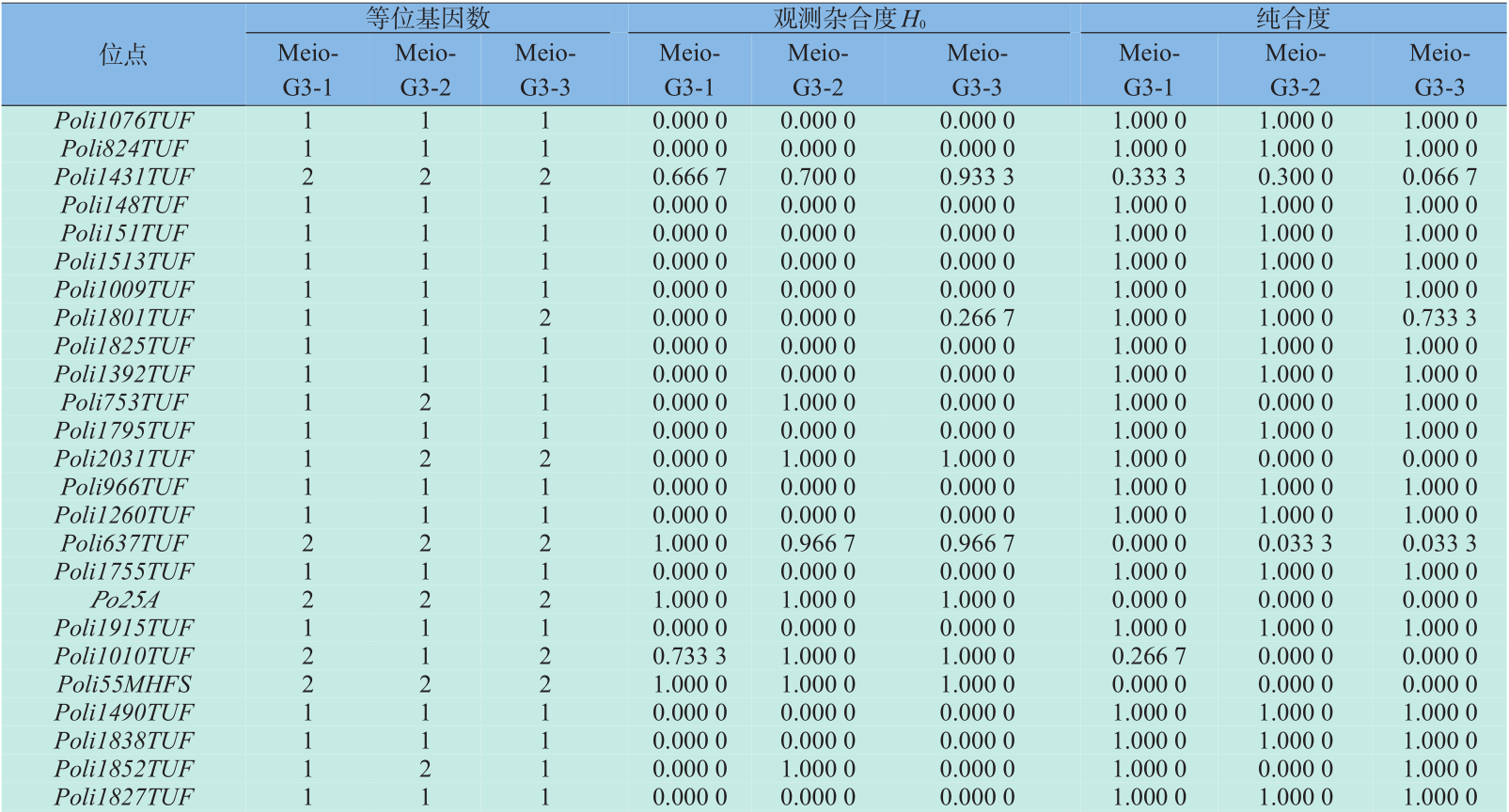
本研究选用的30对微卫星引物在Meio-G3-1、 Meio-G3-2、Meio-G3-3三个家系中均能扩增出稳定的PCR产物,所扩增到的等位基因数分别为35、38、 37,平均等位基因数为1.17、1.27、1.23(见表2)。平均观测杂合度(Ho)分别为0.146 7、0.255 6、0.205 6,平均纯合度分别为0.853 3、0.744 4、0.794 4。 t 检验结果显示,Meio-G3-1、Meio-G3-2、Meio-G3-3 三个家系间的平均观测杂合度和平均纯合度差异均不显著(P >0.05)。
《表2》
表2 30个微卫星座位上的等位基因数、观测杂合度及纯合度
Table 2 Number of alleles,observed heterozygosity and homozygosity for the 30 microsatellite loci

《3.2 连续三代减数分裂雌核发育家系内个体间的遗传相似度》
3.2 连续三代减数分裂雌核发育家系内个体间的遗传相似度
根据所检测的30个微卫星位点的基因类型,分别计算了Meio-G3-1、Meio-G3-2、Meio-G3-3家系内个体间的遗传相似度(见表3)。结果表明,连续三代雌核发育家系内个体间平均遗传相似度在 0.983 8~0.991 8,显示此群体已经是一个遗传一致性很高的品系。另外,各家系母本与子代之间的平均遗传相似度要高于子代之间的相似度。
《表3》
表3 连续三代减数分裂雌核发育家系内的遗传相似度
Table 3 Genetic similarity within family of the third successive meiogynogenetic generation

《3.3 连续三代减数分裂雌核发育家系间的遗传相似度》
3.3 连续三代减数分裂雌核发育家系间的遗传相似度
根据 30 个微卫星位点的基因型数据,计算了 Meio-G3-1、Meio-G3-2、Meio-G3-3三个家系间的遗传相似度(见表 4)。三个家系间的遗传相似度在 0.971 4~0.981 0,遗传距离在0.019 1~0.029 0。三个家系之间均表现出高度的遗传相似性。
《表4》
表4 三个连续三代减数分裂雌核发育家系间的遗传相似系数和遗传距离
Table 4 Genetic similarity and genetic distance among three third-generation of successive meiogynogenetic families in Japanese flounder Paralichthys olivaceus

《3.4 不同微卫星位点的纯合效率和近交系数》
3.4 不同微卫星位点的纯合效率和近交系数
在三个家系中,30 个微卫星位点分别检测出 5 个、8个和7个多态性位点,多态性位点百分比为16.67、26.67、23.33,其余的各个位点,母本检测到的为单态,只有一个等位基因,子代在这些位点的纯合性和母本相同,完全纯合固定。近交系数是衡量微卫星位点纯合速率的一个指标,杂合位点在各家系的近交系数范围为 0~0.7(Meio-G3-1)、0~0.66(Meio-G3-2)、0~0.98(Meio-G3-3)。不同杂合位点在不同家系表现出不同的纯合速率。比如,Poli1431TUF 位点在 Meio-G3-1 和 Meio-G3-2 家系各观测到10个和9个纯合个体,而在Meio-G3-3家系中,只有两个个体在此位点为纯合;Poli1010TUF位点在 Meio-G3-1 家系中观测到 8 个纯合个体,而在 Meio-G3-2和Meio-G3-3家系中却没有观测到纯合个体(见表5)。
《表5》
表5 部分微卫星位点上纯合个体数量、比率、重组率和近交系数
Table 5 Number of homozygote,recombination frequency,coefficient of inbreeding (F) of some detected microsatellite loci in the third successive meiogynogenetic generation offspring Paralichthys olivaceus

《4 结语》
4 结语
本研究首次报道了牙鲆三个连续三代减数分裂雌核发育家系的建立,并通过30对微卫星引物对这三个家系进行了遗传特征的研究。结果表明,家系内个体间和家系间都具有高度的遗传相似性。本研究中只选取一尾亲鱼的卵子,通过人工诱导减数分裂雌核发育,获得减数分裂雌核发育一代家系 Meio-G1,再随机选取 Meio-G1 内一尾发育成熟雌性个体进行第二轮减数分裂雌核发育的诱导,获得第二代减数分裂雌核发育家系Meio-G2。待MeioG2家系雌鱼性成熟时,再随机选取三尾分别进行第三轮减数分裂雌核发育的诱导,从而建立了三个连续三代减数分裂雌核发育家系。每一代都只选用单一母本分别进行减数分裂雌核发育的诱导,在最大程度上确保了遗传背景的一致性和等位基因代际纯化的最大化,从而获得遗传相似度高的家系。
遗传一致性高的动物品系对动物实验的可靠性以及可重复性具有重要意义。在生物学研究中,获得的实验结果包含了处理效应、所用生物的遗传变异、非遗传变异以及各因素之间的互作。一个好的实验设计是要控制和减少这些变异,从而让处理的信息更加清晰和可靠。获得具有高统计效力实验结果的最佳方法是利用不同近交系具有遗传异质性的少量个体进行实验[10] 。在动物中,建立近交系最常用的方法为全同胞交配法,运用此方法,建立一个近交系需要至少20代[11] 。迄今为止,运用全同胞交配法,已经建立了数百个啮齿类的近交系[12] 。在斑马鱼上,一个连续16代的全同胞交配品系也已经被建立[13] 。相比于传统的全同胞交配法,人工诱导减数分裂雌核发育法具有快速固定母本性状、提高等位基因纯合速率等优点。一般认为,一代雌核发育相当于 8~10 代的连续全同胞交配 [14] 。在牙鲆上,一代减数分裂雌核发育相当于 9~10 个世代的全同胞交配,连续第二代减数分裂相当于11~12代全同胞交配[7] 。本研究中,三个家系内子代间的平均遗传相似度在 0.981 3~0.991 8,家系间的平均遗传相似度 0.971 4~0.981 0。团头鲂(Megalobrama amblycephala)连续三代减数分裂雌核发育家系内个体间的平均遗传相似度为 0.984 5[15] ,本研究结果与之相近。通过诱导连续三代雌核发育所获得的个体,无论家系内,还是家系间,都具有高度的遗传相似性,接近或略高于连续 20代全同胞交配所获得的理论结果(0.986)。因此,连续诱导多代减数分裂雌核发育能建立近交系,且比传统的全同胞交配法更节省时间。
由于减数分裂过程中,存在着等位基因的重组,而且由于不同等位基因在染色体上的位置不同造成重组率的差异,导致了纯合速率的差异,所以导致连续多代减数分裂雌核发育虽能一定程度加速等位基因纯化的进程,但很难获得所有位点全部纯合的个体。将连续多代减数分裂雌核发育个体再进行一次有丝分裂雌核发育的诱导,将能获得双单倍体纯合子。在牙鲆上,纯合度高的个体比纯合度低的个体具有更高的有丝分裂雌核发育诱导成功率。
综上所述,通过连续三代减数分裂雌核发育诱导所获得的牙鲆,具有高度的纯合性和遗传相似度,是进行牙鲆良种选育(quantitative trait locus)、 QTL定位及其他遗传学研究的优质材料。











 京公网安备 11010502051620号
京公网安备 11010502051620号




